Serotonina
 | |
| Datos clínicos | |
|---|---|
| Otros nombres | 5-HT, 5-hidroxitriptamina, enteramina, trombocitina, solución de 3-(β-aminoetil)-5-hidroxilo, trombotonina |
| Datos fisiológicos | |
| Tejidos de origen | núcleos del rafe , células enterocromafines |
| Tejidos diana | Todo el sistema |
| Receptores | 5-HT 1 , 5-HT 2 , 5-HT 3 , 5-HT 4 , 5-HT 5 , 5-HT 6 , 5-HT 7 |
| Agonistas | Indirectamente: ISRS , IMAO |
| Precursor | 5-HTP |
| Biosíntesis | Descarboxilasa de L -aminoácidos aromáticos |
| Metabolismo | MAO |
| Identificadores | |
| |
| Número CAS |
|
| Identificador de centro de PubChem |
|
| Unión Internacional para la Investigación y el Desarrollo (IUPHAR)/BPS |
|
| Araña química |
|
| BARRIL |
|
| Ligando PDB |
|
| Panel de control CompTox ( EPA ) |
|
| Tarjeta informativa de la ECHA | 100.000.054 |
 | |
| Nombres | |
|---|---|
| Nombre IUPAC 5-Hidroxitriptamina | |
| Nombre IUPAC preferido 3-(2-aminoetil)-1H - indol-5-ol | |
| Otros nombres 5-Hidroxitriptamina, 5-HT, Enteramina; Trombocitina, 3-(β-Aminoetil)-5-hidroxiindol, 3-(2-Aminoetil)indol-5-ol, Trombotonina | |
| Identificadores | |
| |
Modelo 3D ( JSmol ) |
|
| EBICh |
|
| Química biológica |
|
| Araña química |
|
| Tarjeta informativa de la ECHA | 100.000.054 |
| |
| BARRIL |
|
| Malla | Serotonina |
Identificador de centro de PubChem |
|
| UNIVERSIDAD |
|
Panel de control CompTox ( EPA ) |
|
| |
| |
| Propiedades | |
| C10H12N2O | |
| Masa molar | 176,215 g/mol |
| Apariencia | Polvo blanco |
| Punto de fusión | 167,7 °C (333,9 °F; 440,8 K) 121–122 °C (ligroína) [3] |
| Punto de ebullición | 416 ± 30 °C (a 760 Torr) [1] |
| ligeramente soluble | |
| Acidez (p K a ) | 10,16 en agua a 23,5 °C [2] |
| 2,98 dólares | |
| Peligros | |
| Dosis o concentración letal (LD, LC): | |
LD 50 ( dosis media ) | 750 mg/kg (subcutánea, rata), [4] 4500 mg/kg (intraperitoneal, rata), [5] 60 mg/kg (oral, rata) |
| Ficha de datos de seguridad (FDS) | Hoja de datos de seguridad externa |
Salvo que se indique lo contrario, los datos se proporcionan para los materiales en su estado estándar (a 25 °C [77 °F], 100 kPa). | |


La serotonina ( / ˌsɛrəˈt oʊnɪn , ˌsɪərə- / ) [ 6] [7] [8] o 5 - hidroxitriptamina ( 5-HT ) es un neurotransmisor monoamínico . Su función biológica es compleja y afecta a diversas funciones, como el estado de ánimo , la cognición , la recompensa , el aprendizaje , la memoria y numerosos procesos fisiológicos como los vómitos y la vasoconstricción . [9]
La serotonina se produce en el sistema nervioso central (SNC), específicamente en los núcleos del rafe del tronco encefálico , las células de Merkel de la piel , las células neuroendocrinas pulmonares y las células receptoras del gusto de la lengua . Aproximadamente el 90% de la serotonina que produce el cuerpo humano se encuentra en las células enterocromafines del tracto gastrointestinal , donde regula los movimientos intestinales. [10] [11] [12] Además, se almacena en las plaquetas sanguíneas y se libera durante la agitación y la vasoconstricción, donde luego actúa como agonista de otras plaquetas. [13] Aproximadamente el 8% se encuentra en las plaquetas y el 1–2% en el SNC. [14]
La serotonina se secreta luminal y basolateralmente , lo que conduce a una mayor captación de serotonina por las plaquetas circulantes y a su activación tras la estimulación, lo que aumenta la estimulación de las neuronas mientéricas y la motilidad gastrointestinal . [15] El resto se sintetiza en las neuronas serotoninérgicas del SNC, donde tiene diversas funciones, entre ellas la regulación del estado de ánimo, el apetito y el sueño . [16] [ fuente médica no confiable ] [17] [ fuente médica no confiable ]
La serotonina secretada por las células enterocromafines finalmente encuentra su camino fuera de los tejidos hacia la sangre. Allí, es absorbida activamente por las plaquetas sanguíneas, que la almacenan. Cuando las plaquetas se unen a un coágulo , liberan serotonina, donde puede servir como vasoconstrictor o vasodilatador mientras regula la hemostasia y la coagulación sanguínea. En altas concentraciones, la serotonina actúa como vasoconstrictor al contraer el músculo liso endotelial directamente o al potenciar los efectos de otros vasoconstrictores (por ejemplo, angiotensina II y norepinefrina). La propiedad vasoconstrictora se observa principalmente en estados patológicos que afectan al endotelio, como la aterosclerosis o la hipertensión crónica . En estados fisiológicos normales, la vasodilatación ocurre a través de la liberación mediada por serotonina de óxido nítrico de las células endoteliales y la inhibición de la liberación de norepinefrina de los nervios adrenérgicos . [18] La serotonina también es un factor de crecimiento para algunos tipos de células, lo que puede darle un papel en la cicatrización de heridas. Existen varios receptores de serotonina .
Bioquímicamente, la molécula de indolamina deriva del aminoácido triptófano . La serotonina se metaboliza principalmente a ácido 5-hidroxiindolacético (5-HIAA), principalmente en el hígado . Varias clases de antidepresivos , como los inhibidores selectivos de la recaptación de serotonina (ISRS) y los inhibidores de la recaptación de serotonina y noradrenalina (IRSN), interfieren con la reabsorción normal de serotonina una vez que se realiza la transmisión de la señal, aumentando así los niveles de neurotransmisores en las sinapsis .
Además de los mamíferos, la serotonina se encuentra en todos los animales bilaterales , incluidos gusanos e insectos, [19] así como en hongos y plantas . [20] La presencia de serotonina en venenos de insectos y espinas de plantas sirve para causar dolor, que es un efecto secundario de la inyección de serotonina. [21] [22] La serotonina es producida por amebas patógenas, causando diarrea en el intestino humano. [23] Su presencia generalizada en muchas semillas y frutas puede servir para estimular el tracto digestivo para que expulse las semillas. [24] [ verificación fallida ]
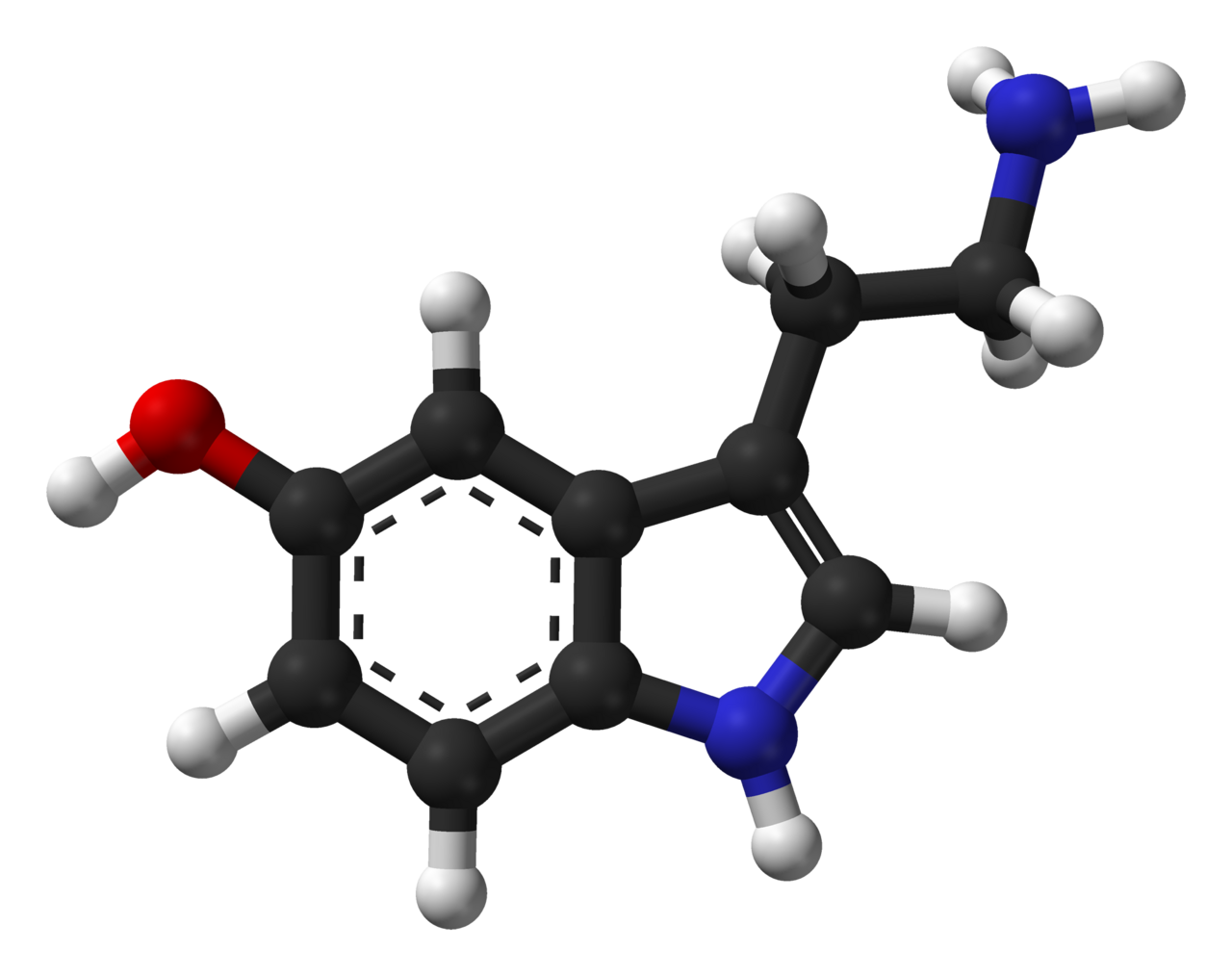
Estructura molecular
Bioquímicamente, la molécula de indolamina deriva del aminoácido triptófano , a través de la hidroxilación (limitante de la velocidad) de la posición 5 del anillo (formando el intermediario 5-hidroxitriptófano ), y luego la descarboxilación para producir serotonina. [25] Las conformaciones preferibles se definen a través de la cadena de etilamina, lo que da como resultado seis conformaciones diferentes. [26]
Estructura cristalina
La serotonina cristaliza en el grupo espacial quiral P2 1 2 1 2 1 formando diferentes interacciones de enlaces de hidrógeno entre las moléculas de serotonina a través de enlaces intermoleculares NH...O y OH...N. [27] La serotonina también forma varias sales, incluida la formulación farmacéutica de adipato de serotonina. [28]
Papel biológico
La serotonina está involucrada en numerosos procesos fisiológicos, [29] incluyendo el sueño , [30] la termorregulación , el aprendizaje y la memoria , el dolor , el comportamiento (social), [31] la actividad sexual , la alimentación, la actividad motora, el desarrollo neuronal, [32] y los ritmos biológicos . [33] En animales menos complejos, como algunos invertebrados , la serotonina regula la alimentación y otros procesos. [34] En las plantas la síntesis de serotonina parece estar asociada con señales de estrés. [20] [35] A pesar de su prominencia de larga data en la publicidad farmacéutica, la afirmación de que los niveles bajos de serotonina causan depresión no está respaldada por evidencia científica. [36] [37] [38]
Efectos celulares
La serotonina actúa principalmente a través de sus receptores y sus efectos dependen de qué células y tejidos expresan estos receptores. [33]
El metabolismo implica primero la oxidación por la monoaminooxidasa a 5-hidroxiindolacetaldehído (5-HIAL). [39] [40] El paso limitante es la transferencia de hidruro de serotonina al cofactor flavina. [41] A continuación, la oxidación por la aldehído deshidrogenasa (ALDH) a ácido 5-hidroxiindolacético ( 5-HIAA ), el derivado del ácido indol acético. Este último luego se excreta por los riñones.
Receptores
Los receptores 5-HT, los receptores de serotonina, se encuentran en la membrana celular de las células nerviosas y otros tipos de células en animales, y median los efectos de la serotonina como ligando endógeno y de una amplia gama de fármacos y drogas psicodélicas . A excepción del receptor 5-HT 3 , un canal iónico controlado por ligando , todos los demás receptores 5-HT son receptores acoplados a proteína G (también llamados receptores de siete transmembrana o heptahelicoidales) que activan una cascada de segundos mensajeros intracelulares . [42]
Terminación
La acción serotoninérgica termina principalmente a través de la captación de 5-HT desde la sinapsis. Esto se logra a través del transportador de monoamina específico para 5-HT, SERT , en la neurona presináptica. Varios agentes pueden inhibir la recaptación de 5-HT, incluyendo la cocaína , el dextrometorfano (un antitusivo ), los antidepresivos tricíclicos y los inhibidores selectivos de la recaptación de serotonina (ISRS). Un estudio de 2006 encontró que una parte significativa de la depuración sináptica de 5-HT se debe a la actividad selectiva del transportador de monoamina de la membrana plasmática (PMAT) que transporta activamente la molécula a través de la membrana y de regreso a la célula presináptica. [43]
En contraste con la alta afinidad de SERT, el PMAT ha sido identificado como un transportador de baja afinidad, con una Km aparente de 114 micromoles/l para la serotonina, que es aproximadamente 230 veces mayor que la de SERT. Sin embargo, el PMAT, a pesar de su afinidad serotoninérgica relativamente baja, tiene una "capacidad" de transporte considerablemente mayor que SERT, "lo que resulta en eficiencias de captación aproximadamente comparables a SERT ... en sistemas de expresión heterólogos". [43] El estudio también sugiere que la administración de ISRS como fluoxetina y sertralina puede estar asociada con un efecto inhibidor sobre la actividad de PMAT cuando se utilizan en dosis más altas que las normales ( los valores de la prueba IC50 utilizados en los ensayos fueron 3-4 veces más altos que la dosis prescriptiva típica).
Serotonilación
La serotonina también puede enviar señales a través de un mecanismo no receptor llamado serotonina, en el que la serotonina modifica las proteínas. [44] Este proceso es la base de los efectos de la serotonina sobre las células formadoras de plaquetas ( trombocitos ), en las que se vincula con la modificación de las enzimas de señalización llamadas GTPasas , que luego desencadenan la liberación del contenido de las vesículas por exocitosis . [45] Un proceso similar es la base de la liberación pancreática de insulina. [44]
Los efectos de la serotonina sobre el tono del músculo liso vascular (la función biológica que le dio nombre originalmente a la serotonina) dependen de la serotonina de las proteínas implicadas en el aparato contráctil de las células musculares. [46]
| Receptor | Ki (nM) [47 ] | Función del receptor [Nota 1] |
|---|---|---|
| La familia de receptores 5-HT 1 envía señales a través de la inhibición de G i/o de la adenilil ciclasa . | ||
| 5-HT 1A | 3.17 | Memoria [ vaga ] (agonistas ↓); aprendizaje [ vaga ] (agonistas ↓); ansiedad (agonistas ↓); depresión (agonistas ↓); síntomas positivos, negativos y cognitivos de la esquizofrenia (agonistas parciales ↓); analgesia (agonistas ↑); agresión (agonistas ↓); liberación de dopamina en la corteza prefrontal (agonistas ↑); liberación y síntesis de serotonina (agonistas ↓) |
| 5-HT 1B | 4.32 | Vasoconstricción (agonistas ↑); agresión (agonistas ↓); masa ósea (↓). Autorreceptor de serotonina. |
| 5-HT 1D | 5.03 | Vasoconstricción (agonistas ↑) |
| 5-HT 1E | 7.53 | |
| 5-HT 1F | 10 | |
| La familia de receptores 5-HT 2 envía señales a través de la activación de la fosfolipasa C por G q . | ||
| 5- HT2A | 11.55 | Psicodelia (agonistas ↑); depresión (agonistas y antagonistas ↓); ansiedad (antagonistas ↓); síntomas positivos y negativos de la esquizofrenia (antagonistas ↓); liberación de noradrenalina del locus coeruleus (antagonistas ↑); liberación de glutamato en la corteza prefrontal (agonistas ↑); dopamina en la corteza prefrontal (agonistas ↑); [48] contracciones de la vejiga urinaria (agonistas ↑) [49] |
| 5- HT2B | 8.71 | Funcionamiento cardiovascular (los agonistas aumentan el riesgo de hipertensión pulmonar), empatía (a través de las neuronas de von Economo [50] ) |
| 5- HT2C | 5.02 | Liberación de dopamina en la vía mesocorticolímbica (agonistas ↓); liberación de acetilcolina en la corteza prefrontal (agonistas ↑); actividad dopaminérgica y noradrenérgica en la corteza frontal (antagonistas ↑); [51] apetito (agonistas ↓); efectos antipsicóticos (agonistas ↑); efectos antidepresivos (agonistas y antagonistas ↑) |
| Otros receptores 5-HT | ||
| 5- HT3 | 593 | Emesis (agonistas ↑); ansiolisis (antagonistas ↑). |
| 5- HT4 | 125,89 | Movimiento de los alimentos a través del tracto gastrointestinal (agonistas ↑); memoria y aprendizaje (agonistas ↑); efectos antidepresivos (agonistas ↑). Señalización a través de la activación de la adenilato ciclasa por G αs . |
| 5-HT 5A | 251.2 | Consolidación de la memoria. [52] Señales vía G i/o inhibición de la adenilil ciclasa . |
| 5- HT6 | 98,41 | Cognición (antagonistas ↑); efectos antidepresivos (agonistas y antagonistas ↑); efectos ansiogénicos (antagonistas ↑ [53] ). Señalización de G s a través de la activación de la adenilil ciclasa . |
| 5- HT7 | 8.11 | Cognición (antagonistas ↑); efectos antidepresivos (antagonistas ↑). Actúa mediante la señalización de G s mediante la activación de la adenilil ciclasa . |
Sistema nervioso

Las neuronas de los núcleos del rafe son la principal fuente de liberación de 5-HT en el cerebro. [54] Hay nueve núcleos del rafe, designados B1–B9, que contienen la mayoría de las neuronas que contienen serotonina (algunos científicos eligieron agrupar los núcleos rafes lineares en un solo núcleo), todos los cuales están ubicados a lo largo de la línea media del tronco encefálico y centrados en la formación reticular . [55] [56] Los axones de las neuronas de los núcleos del rafe forman un sistema de neurotransmisores que llega a casi todas las partes del sistema nervioso central. Los axones de las neuronas en los núcleos inferiores del rafe terminan en el cerebelo y la médula espinal , mientras que los axones de los núcleos superiores se extienden por todo el cerebro.
Ultraestructura y función
Los núcleos de serotonina también pueden dividirse en dos grupos principales, el rostral y el caudal, que contienen tres y cuatro núcleos respectivamente. El grupo rostral consta de los núcleos lineales caudales (B8), los núcleos del rafe dorsal (B6 y B7) y los núcleos del rafe medio (B5, B8 y B9), que se proyectan hacia múltiples estructuras corticales y subcorticales. El grupo caudal consta del núcleo magno del rafe (B3), el núcleo oscuro del rafe (B2), el núcleo pálido del rafe (B1) y la formación reticular medular lateral, que se proyectan hacia el tronco encefálico. [57]
La vía serotoninérgica está involucrada en la función sensoriomotora, con vías que se proyectan tanto en áreas corticales (núcleos del rafe dorsal y medio), subcorticales y espinales involucradas en la actividad motora. La manipulación farmacológica sugiere que la actividad serotoninérgica aumenta con la actividad motora mientras que las tasas de activación de las neuronas serotoninérgicas aumentan con estímulos visuales intensos. Los modelos animales sugieren que la señalización del kainato regula negativamente las acciones de la serotonina en la retina, con posibles implicaciones para el control del sistema visual. [58] Las proyecciones descendentes forman una vía que inhibe el dolor llamada "vía inhibitoria descendente" que puede ser relevante para un trastorno como la fibromialgia, la migraña y otros trastornos del dolor, y la eficacia de los antidepresivos en ellos. [59]
Las proyecciones serotoninérgicas de los núcleos caudales están involucradas en la regulación del estado de ánimo y la emoción, y los estados hipo- [60] o hiperserotoninérgicos [61] pueden estar involucrados en la depresión y la conducta enferma.
Microanatomía
La serotonina se libera en la sinapsis, o espacio entre neuronas, y se difunde en un espacio relativamente amplio (>20 nm) para activar los receptores 5-HT ubicados en las dendritas , los cuerpos celulares y las terminales presinápticas de las neuronas adyacentes.
Cuando los humanos huelen comida, se libera dopamina para aumentar el apetito . Pero, a diferencia de lo que ocurre en los gusanos, la serotonina no aumenta el comportamiento anticipatorio en los humanos; en cambio, la serotonina liberada mientras se consume activa los receptores 5-HT2C en las células productoras de dopamina. Esto detiene su liberación de dopamina y, por lo tanto, la serotonina disminuye el apetito. Los medicamentos que bloquean los receptores 5-HT 2C hacen que el cuerpo sea incapaz de reconocer cuándo ya no tiene hambre o necesita nutrientes, y se asocian con el aumento de peso, [62] especialmente en personas con un bajo número de receptores. [63] La expresión de los receptores 5-HT 2C en el hipocampo sigue un ritmo diurno , [64] al igual que la liberación de serotonina en el núcleo ventromedial , que se caracteriza por un pico en la mañana cuando la motivación para comer es más fuerte. [65]
En los macacos , los machos alfa tienen el doble de niveles de serotonina en el cerebro que los machos y hembras subordinados (medido por la concentración de 5-HIAA en el líquido cefalorraquídeo (LCR)). El estado de dominancia y los niveles de serotonina en el LCR parecen estar correlacionados positivamente. Cuando los machos dominantes fueron eliminados de dichos grupos, los machos subordinados comenzaron a competir por el dominio. Una vez que se establecieron nuevas jerarquías de dominancia, los niveles de serotonina de los nuevos individuos dominantes también aumentaron hasta duplicar los de los machos y hembras subordinados. Aún no se ha establecido la razón por la que los niveles de serotonina solo son altos en los machos dominantes, pero no en las hembras dominantes. [66]
En los seres humanos, los niveles de inhibición del receptor 5-HT 1A en el cerebro muestran una correlación negativa con la agresión [67], y una mutación en el gen que codifica el receptor 5-HT 2A puede duplicar el riesgo de suicidio en aquellos con ese genotipo [68] . La serotonina en el cerebro no suele degradarse después de su uso, sino que es recogida por las neuronas serotoninérgicas mediante transportadores de serotonina en sus superficies celulares. Los estudios han revelado que casi el 10% de la varianza total en la personalidad relacionada con la ansiedad depende de variaciones en la descripción de dónde, cuándo y cuántos transportadores de serotonina deben desplegar las neuronas [69] .
Fuera del sistema nervioso
Tracto digestivo (emético)
La serotonina regula la función gastrointestinal (GI). El intestino está rodeado de células enterocromafines , que liberan serotonina en respuesta a los alimentos en el lumen . Esto hace que el intestino se contraiga alrededor de los alimentos. Las plaquetas en las venas que drenan el intestino recogen el exceso de serotonina. A menudo hay anomalías de serotonina en trastornos gastrointestinales como el estreñimiento y el síndrome del intestino irritable. [70]
Si hay irritantes presentes en el alimento, las células enterocromafines liberan más serotonina para hacer que el intestino se mueva más rápido, es decir, para provocar diarrea, por lo que el intestino se vacía de la sustancia nociva. Si la serotonina se libera en la sangre más rápido de lo que las plaquetas pueden absorberla, el nivel de serotonina libre en la sangre aumenta. Esto activa los receptores 5-HT3 en la zona gatillo de los quimiorreceptores que estimulan el vómito . [71] Así, los fármacos y las toxinas estimulan la liberación de serotonina de las células enterocromafines en la pared intestinal. Las células enterocromafines no sólo reaccionan a los malos alimentos, sino que también son muy sensibles a la irradiación y a la quimioterapia contra el cáncer . Los fármacos que bloquean la 5HT3 son muy eficaces para controlar las náuseas y los vómitos producidos por el tratamiento del cáncer, y se consideran el estándar de oro para este propósito. [72]
Pulmones
El pulmón , [73] incluido el de los reptiles, [74] contiene células epiteliales especializadas que se presentan como células solitarias o como grupos llamados cuerpos neuroepiteliales o células bronquiales de Kulchitsky o alternativamente células K. [75] Estas son células enterocromafines que, al igual que las del intestino, liberan serotonina. [ 75] Su función es probablemente la vasoconstricción durante la hipoxia . [73]
Piel
La serotonina también es producida por las células de Merkel , que forman parte del sistema somatosensorial . [76]
Metabolismo óseo
En ratones y humanos, se ha demostrado que las alteraciones en los niveles de serotonina y la señalización regulan la masa ósea. [77] [78] [79] [80] Los ratones que carecen de serotonina cerebral tienen osteopenia , mientras que los ratones que carecen de serotonina intestinal tienen alta densidad ósea. En humanos, se ha demostrado que el aumento de los niveles de serotonina en sangre es un predictor negativo significativo de baja densidad ósea. La serotonina también se puede sintetizar, aunque a niveles muy bajos, en las células óseas. Media sus acciones en las células óseas utilizando tres receptores diferentes. A través de los receptores 5-HT 1B , regula negativamente la masa ósea, mientras que lo hace positivamente a través de los receptores 5-HT 2B y los receptores 5-HT 2C . Existe un equilibrio muy delicado entre el papel fisiológico de la serotonina intestinal y su patología. El aumento del contenido extracelular de serotonina da como resultado una retransmisión compleja de señales en los osteoblastos que culmina en eventos transcripcionales dependientes de FoxO1/Creb y ATF4. [81] Tras los hallazgos de 2008 de que la serotonina intestinal regula la masa ósea, han comenzado las investigaciones mecanicistas sobre lo que regula la síntesis de serotonina del intestino en la regulación de la masa ósea. Se ha demostrado que Piezo1 detecta el ARN en el intestino y transmite esta información a través de la síntesis de serotonina al hueso actuando como un sensor de ARN monocatenario (ssRNA) que regula la producción de 5-HT. La eliminación específica del epitelio intestinal de Piezo1 de ratón alteró profundamente la peristalsis intestinal, impidió la colitis experimental y suprimió los niveles séricos de 5-HT. Debido a la deficiencia sistémica de 5-HT, la eliminación condicional de Piezo1 aumentó la formación ósea. En particular, el ssRNA fecal se identificó como un ligando natural de Piezo1, y la síntesis de 5-HT estimulada por ssRNA del intestino se evocó de una manera independiente de MyD88/TRIF. La infusión colónica de ARNasa A suprimió la motilidad intestinal y aumentó la masa ósea. Estos hallazgos sugieren que el ARN monocatenario intestinal es un determinante principal de los niveles sistémicos de 5-HT, lo que indica que el eje ARN monocatenario-Piezo1 es un objetivo profiláctico potencial para el tratamiento de trastornos óseos e intestinales. Los estudios realizados en 2008, 2010 y 2019 han abierto la posibilidad de que la investigación sobre la serotonina trate los trastornos de la masa ósea. [82] [83]
Desarrollo de órganos
Dado que la serotonina indica la disponibilidad de recursos, no es sorprendente que afecte al desarrollo de los órganos. Muchos estudios en humanos y animales han demostrado que la nutrición en la vida temprana puede influir, en la edad adulta, en aspectos como la grasa corporal, los lípidos en sangre, la presión arterial, la aterosclerosis , el comportamiento, el aprendizaje y la longevidad. [84] [85] [86] Un experimento con roedores muestra que la exposición neonatal a los ISRS produce cambios persistentes en la transmisión serotoninérgica del cerebro que resultan en cambios de comportamiento, [87] [88] que se revierten con el tratamiento con antidepresivos. [89] Al tratar con fluoxetina a ratones normales y knock out que carecían del transportador de serotonina, los científicos demostraron que las reacciones emocionales normales en la edad adulta, como una latencia corta para escapar de las descargas eléctricas en las patas y la inclinación a explorar nuevos entornos, dependían de transportadores de serotonina activos durante el período neonatal. [90] [91]
La serotonina humana también puede actuar directamente como factor de crecimiento . El daño hepático aumenta la expresión celular de los receptores 5-HT 2A y 5-HT 2B , lo que media el recrecimiento compensatorio del hígado (véase Hígado § Regeneración y trasplante ) [92]. La serotonina presente en la sangre estimula entonces el crecimiento celular para reparar el daño hepático. [93]
Los receptores 5-HT 2B también activan los osteocitos , que forman el hueso [94] Sin embargo, la serotonina también inhibe a los osteoblastos , a través de los receptores 5-HT 1B . [95]
Factor de crecimiento cardiovascular
Además, la serotonina provoca la activación de la óxido nítrico sintasa endotelial y estimula, a través de un mecanismo mediado por el receptor 5-HT1B , la fosforilación de la activación de la proteína quinasa activada por mitógeno p44/p42 en cultivos de células endoteliales aórticas bovinas. [ aclaración necesaria ] [96] En la sangre, la serotonina es recogida del plasma por las plaquetas, que la almacenan. Por lo tanto, es activa dondequiera que las plaquetas se unan en el tejido dañado, como vasoconstrictor para detener el sangrado, y también como un factor mitótico de fibrocitos (factor de crecimiento), para ayudar a la curación. [97]
Farmacología
Varias clases de medicamentos tienen como objetivo el sistema de serotonina, incluidos algunos antidepresivos , antipsicóticos , ansiolíticos , antieméticos y antimigrañosos , así como también los medicamentos psicodélicos y los entactógenos .
Mecanismo de acción
En reposo, la serotonina se almacena en las vesículas de las neuronas presinápticas. Cuando se estimula mediante impulsos nerviosos, la serotonina se libera como neurotransmisor en la sinapsis, uniéndose reversiblemente al receptor postsináptico para inducir un impulso nervioso en la neurona postsináptica. La serotonina también puede unirse a los autorreceptores de la neurona presináptica para regular la síntesis y liberación de serotonina. Normalmente, la serotonina vuelve a la neurona presináptica para detener su acción y luego se reutiliza o se descompone por acción de la monoaminooxidasa. [98]
Drogas psicodélicas
Las drogas psicodélicas serotoninérgicas psilocina / psilocibina , DMT , mescalina , hongos psicodélicos y LSD son agonistas , principalmente en los receptores 5-HT 2A y 5-HT 2C . [99] [100] [101] El empatógeno-entactógeno MDMA libera serotonina de las vesículas sinápticas de las neuronas. [102]
Antidepresivos
Los fármacos que alteran los niveles de serotonina se utilizan para tratar la depresión , el trastorno de ansiedad generalizada y la fobia social . Los inhibidores de la monoaminooxidasa (IMAO) impiden la descomposición de los neurotransmisores monoamínicos (incluida la serotonina) y, por lo tanto, aumentan las concentraciones del neurotransmisor en el cerebro. La terapia con IMAO se asocia con muchas reacciones adversas a los medicamentos y los pacientes corren el riesgo de sufrir una emergencia hipertensiva desencadenada por alimentos con alto contenido de tiramina y ciertos medicamentos. Algunos medicamentos inhiben la recaptación de serotonina, lo que hace que permanezca en la hendidura sináptica durante más tiempo. Los antidepresivos tricíclicos (ATC) inhiben la recaptación tanto de serotonina como de noradrenalina . Los inhibidores selectivos de la recaptación de serotonina ( ISRS ) más nuevos tienen menos efectos secundarios y menos interacciones con otros medicamentos. [103]
Se ha demostrado que ciertos medicamentos ISRS reducen los niveles de serotonina por debajo del valor basal después del uso crónico, a pesar de los aumentos iniciales. [104] El gen 5-HTTLPR codifica la cantidad de transportadores de serotonina en el cerebro, y una mayor cantidad de transportadores de serotonina causa una disminución en la duración y magnitud de la señalización serotoninérgica. [105] También se ha descubierto que el polimorfismo 5-HTTLPR (l/l) que hace que se formen más transportadores de serotonina es más resistente contra la depresión y la ansiedad. [106] [107]
Síndrome serotoninérgico
Los niveles extremadamente altos de serotonina pueden causar una afección conocida como síndrome serotoninérgico , con efectos tóxicos y potencialmente fatales. En la práctica, tales niveles tóxicos son esencialmente imposibles de alcanzar mediante una sobredosis de un solo fármaco antidepresivo, sino que requieren una combinación de agentes serotoninérgicos, como un ISRS con un IMAO , que puede presentarse en dosis terapéuticas. [108] [109] La intensidad de los síntomas del síndrome serotoninérgico varía en un amplio espectro, y las formas más leves se observan incluso en niveles no tóxicos. [110] Se estima que el 14% de los pacientes que experimentan síndrome serotoninérgico sufren una sobredosis de ISRS; mientras tanto, la tasa de mortalidad está entre el 2% y el 12%. [108] [111] [112]
Antieméticos
Algunos antagonistas de 5-HT 3 , como ondansetrón , granisetrón y tropisetrón , son agentes antieméticos importantes . Son particularmente importantes en el tratamiento de las náuseas y los vómitos que se producen durante la quimioterapia contra el cáncer con fármacos citotóxicos . Otra aplicación es en el tratamiento de las náuseas y los vómitos posoperatorios .
Otro
Algunos fármacos agonistas serotoninérgicos causan fibrosis en cualquier parte del cuerpo, en particular el síndrome de fibrosis retroperitoneal , así como fibrosis de la válvula cardíaca . [113]
En el pasado, tres grupos de fármacos serotoninérgicos se han relacionado epidemiológicamente con estos síndromes. Se trata de los fármacos antimigrañosos vasoconstrictores serotoninérgicos ( ergotamina y metisergida ), [113] los fármacos supresores del apetito serotoninérgicos ( fenfluramina , clorfentermina y aminorex ) y ciertos agonistas dopaminérgicos antiparkinsonianos, que también estimulan los receptores serotoninérgicos 5-HT 2B . Entre ellos se encuentran la pergolida y la cabergolina , pero no la lisurida, más específica de la dopamina . [114]
Al igual que con la fenfluramina, algunos de estos fármacos han sido retirados del mercado después de que los grupos que los tomaban mostraran un aumento estadístico de uno o más de los efectos secundarios descritos. Un ejemplo es la pergolida . El uso de este fármaco estaba disminuyendo desde que en 2003 se informó que estaba asociado con la fibrosis cardíaca. [115]
Dos estudios independientes publicados en The New England Journal of Medicine en enero de 2007 implicaron a la pergolida, junto con la cabergolina , en causar enfermedad cardíaca valvular . [116] [117] Como resultado de esto, la FDA retiró la pergolida del mercado de los Estados Unidos en marzo de 2007. [118] (Dado que la cabergolina no está aprobada en los Estados Unidos para la enfermedad de Parkinson, sino para la hiperprolactinemia, el fármaco permanece en el mercado. El tratamiento para la hiperprolactinemia requiere dosis más bajas que para la enfermedad de Parkinson, lo que disminuye el riesgo de enfermedad cardíaca valvular). [119]
Metiltriptaminas y alucinógenos
Varias plantas contienen serotonina junto con una familia de triptaminas relacionadas que están metiladas en los grupos amino (NH 2 ) y (OH) , son N -óxidos o no tienen el grupo OH. Estos compuestos llegan al cerebro, aunque una parte de ellos son metabolizados por enzimas monoaminooxidasas (principalmente MAO-A ) en el hígado. Algunos ejemplos son las plantas del género Anadenanthera que se utilizan en el rapé alucinógeno yopo . Estos compuestos están ampliamente presentes en las hojas de muchas plantas y pueden servir como elementos disuasorios para la ingestión animal. La serotonina se encuentra en varios hongos del género Panaeolus . [120]
Biología comparada y evolución
Organismos unicelulares
La serotonina es utilizada por una variedad de organismos unicelulares para diversos fines. Se ha descubierto que los ISRS son tóxicos para las algas. [121] El parásito gastrointestinal Entamoeba histolytica secreta serotonina, lo que provoca una diarrea secretora sostenida en algunas personas. [23] [122] Se ha descubierto que los pacientes infectados con E. histolytica tienen niveles séricos de serotonina muy elevados, que volvieron a la normalidad después de la resolución de la infección. [123] E. histolytica también responde a la presencia de serotonina volviéndose más virulenta. [124] Esto significa que la secreción de serotonina no solo sirve para aumentar la propagación de las entamoebas al provocar diarrea al huésped, sino que también sirve para coordinar su comportamiento de acuerdo con su densidad de población, un fenómeno conocido como detección de quórum . Fuera del intestino de un huésped, no hay nada que las entamoebas provoquen para liberar serotonina, por lo tanto, la concentración de serotonina es muy baja. La serotonina baja indica a las entamoebas que están fuera de un huésped y se vuelven menos virulentas para conservar energía. Cuando entran en un nuevo huésped, se multiplican en el intestino y se vuelven más virulentos a medida que las células enterocromafines son provocadas por ellos y la concentración de serotonina aumenta.
Plantas y setas comestibles
Al secar las semillas , la producción de serotonina es una forma de deshacerse de la acumulación de amoníaco venenoso . El amoníaco se recoge y se coloca en la parte indólica del L - triptófano , que luego es descarboxilado por la triptófano descarboxilasa para dar triptamina, que luego es hidroxilada por una citocromo P450 monooxigenasa , produciendo serotonina. [125]
Sin embargo, dado que la serotonina es un modulador importante del tracto gastrointestinal, puede producirse en los frutos de las plantas como una forma de acelerar el paso de las semillas a través del tracto digestivo, de la misma manera que muchos laxantes asociados a semillas y frutas bien conocidos. La serotonina se encuentra en hongos , frutas y verduras . Los valores más altos de 25-400 mg/kg se han encontrado en nueces de los géneros nogal ( Juglans ) y nogal americano ( Carya ). Se han encontrado concentraciones de serotonina de 3-30 mg/kg en plátanos , piñas , bananas , kiwis , ciruelas y tomates . Se han encontrado niveles moderados de 0,1-3 mg/kg en una amplia gama de verduras analizadas. [24] [20]
La serotonina es un compuesto del veneno contenido en las ortigas ( Urtica dioica ), donde causa dolor al ser inyectado de la misma manera que su presencia en los venenos de los insectos. [22] También se encuentra de forma natural en Paramuricea clavata , o abanico del Mar Rojo. [126]
Se han encontrado serotonina y triptófano en chocolate con distintos contenidos de cacao. El contenido más alto de serotonina (2,93 μg/g) se encontró en chocolate con 85% de cacao, y el contenido más alto de triptófano (13,27–13,34 μg/g) se encontró en chocolate con 70–85% de cacao. No se encontró el intermediario en la síntesis de triptófano a serotonina, el 5-hidroxitriptófano. [127]
El desarrollo de las raíces en Arabidopsis thaliana es estimulado y modulado por la serotonina, de diversas maneras y en distintas concentraciones. [128]
La serotonina actúa como un químico de defensa de las plantas contra los hongos. Cuando se infecta con podredumbre de corona por Fusarium ( Fusarium pseudograminearum ), el trigo ( Triticum aestivum ) aumenta en gran medida su producción de triptófano para sintetizar nueva serotonina. [129] La función de esto es poco conocida [129] pero el trigo también produce serotonina cuando se infecta con Stagonospora nodorum , en ese caso para retardar la producción de esporas. [130] El cereal modelo Brachypodium distachyon , utilizado como sustituto de investigación para el trigo y otros cereales de producción, también produce serotonina, cumaroil -serotonina y feruloil -serotonina en respuesta a F. graminearum . Esto produce un ligero efecto antimicrobiano . B. distachyon produce más serotonina (y conjugados) en respuesta a F. graminearum productor de deoxinivalenol (DON) que a F. graminearum no productor de DON. [131] Solanum lycopersicum produce muchos conjugados AA , incluidos varios de serotonina, en sus hojas, tallos y raíces en respuesta a la infección por Ralstonia solanacearum . [132]
Invertebrados
La serotonina funciona como un neurotransmisor en el sistema nervioso de la mayoría de los animales.
Nematodos
Por ejemplo, en el gusano redondo Caenorhabditis elegans , que se alimenta de bacterias, la serotonina se libera como una señal en respuesta a eventos positivos, como encontrar una nueva fuente de alimento o en animales machos encontrar una hembra con la que aparearse. [133] Cuando un gusano bien alimentado siente bacterias en su cutícula , se libera dopamina , que lo ralentiza; si está hambriento, también se libera serotonina, que ralentiza aún más al animal. Este mecanismo aumenta la cantidad de tiempo que los animales pasan en presencia de alimento. [134] La serotonina liberada activa los músculos utilizados para alimentarse, mientras que la octopamina los suprime. [135] [136] La serotonina se difunde a las neuronas sensibles a la serotonina, que controlan la percepción del animal de la disponibilidad de nutrientes.
Decápodos
Si se inyecta serotonina a las langostas , se comportan como individuos dominantes, mientras que la octopamina provoca un comportamiento subordinado . [31] Un cangrejo de río asustado puede mover la cola para huir, y el efecto de la serotonina en este comportamiento depende en gran medida del estatus social del animal. La serotonina inhibe la reacción de huida en los subordinados, pero la potencia en los individuos socialmente dominantes o aislados. La razón de esto es que la experiencia social altera la proporción entre los receptores de serotonina (receptores 5-HT) que tienen efectos opuestos en la respuesta de lucha o huida . [ aclaración necesaria ] El efecto de los receptores 5-HT 1 predomina en los animales subordinados, mientras que los receptores 5-HT 2 predominan en los dominantes. [137]
En venenos
La serotonina es un componente común de los venenos de los invertebrados, las glándulas salivales, los tejidos nerviosos y otros tejidos, en moluscos, insectos, crustáceos, escorpiones, varios tipos de gusanos y medusas. [22] Los Rhodnius prolixus adultos ( hematófagos en los vertebrados) secretan lipocalinas en la herida durante la alimentación. En 2003 se demostró que estas lipocalinas secuestran serotonina para prevenir la vasoconstricción (y posiblemente la coagulación) en el huésped. [138]
Insectos
La serotonina se conserva evolutivamente y aparece en todo el reino animal. Se observa en los procesos de los insectos en funciones similares a las del sistema nervioso central humano, como la memoria, el apetito, el sueño y el comportamiento. [139] [19] Algunos circuitos en los cuerpos de los hongos son serotoninérgicos. [140] (Véase el ejemplo específico de Drosophila a continuación, §Dipterans).
Acrídidos
La enjambre de langostas se inicia pero no se mantiene por la serotonina, [141] y la liberación se desencadena por el contacto táctil entre individuos. [142] Esto transforma la preferencia social de la aversión a un estado gregario que permite grupos coherentes. [143] [142] [141] El aprendizaje en moscas y abejas se ve afectado por la presencia de serotonina. [144] [145]
Papel en los insecticidas
Los receptores 5-HT de los insectos tienen secuencias similares a las versiones de los vertebrados, pero se han observado diferencias farmacológicas. La respuesta a los fármacos en los invertebrados ha sido mucho menos caracterizada que la farmacología en los mamíferos y se ha discutido el potencial de los insecticidas selectivos para cada especie. [146]
Himenópteros
Las avispas y los avispones tienen serotonina en su veneno, [147] que causa dolor e inflamación [21] [22] al igual que los escorpiones . [148] [22] Pheidole dentata asume cada vez más tareas en la colonia a medida que envejece, lo que requiere que responda a más y más señales olfativas en el curso de su realización. Se demostró que esta ampliación de la respuesta olfativa va acompañada de un aumento de serotonina y dopamina , pero no de octopamina en 2006. [149]
Dípteros
Si las moscas son alimentadas con serotonina, son más agresivas; las moscas desprovistas de serotonina aún exhiben agresión, pero lo hacen con mucha menos frecuencia. [150] En sus buches juega un papel vital en la motilidad digestiva producida por la contracción. La serotonina que actúa sobre el buche es exógena al propio buche y la investigación de 2012 sugirió que probablemente se originó en el plexo neural de serotonina en el ganglio singanglionar torácico-abdominal. [151] En 2011 se descubrió que un cuerpo de hongo serotoninérgico de Drosophila trabajaba en conjunto con Amnesiac para formar recuerdos. [140] En 2007 se descubrió que la serotonina promovía la agresión en Diptera , que era contrarrestada por el neuropéptido F, un hallazgo sorprendente dado que ambos promueven el cortejo , que suele ser similar a la agresión en la mayoría de los aspectos. [140]
Vertebrados
La serotonina, también conocida como 5-hidroxitriptamina (5-HT), es un neurotransmisor más conocido por su participación en los trastornos del estado de ánimo en los seres humanos. También es un neuromodulador ampliamente presente entre vertebrados e invertebrados. [152] Se ha descubierto que la serotonina tiene asociaciones con muchos sistemas fisiológicos como el cardiovascular, la termorregulación y las funciones conductuales, incluidos: el ritmo circadiano , el apetito, el comportamiento agresivo y sexual, la reactividad sensoriomotora y el aprendizaje, y la sensibilidad al dolor. [153] Se discutirá más a fondo la función de la serotonina en los sistemas neurológicos junto con comportamientos específicos entre vertebrados que se encuentran fuertemente asociados con la serotonina. También se mencionan dos estudios de caso relevantes sobre el desarrollo de la serotonina que involucran peces teleósteos y ratones .
En los mamíferos, la 5-HT se concentra en gran medida en la sustancia negra , el área tegmental ventral y los núcleos del rafe . Otras áreas de menor concentración incluyen otras regiones cerebrales y la médula espinal. [152] También se ha demostrado que las neuronas 5-HT están muy ramificadas, lo que indica que son estructuralmente prominentes para influir en múltiples áreas del SNC al mismo tiempo, aunque esta tendencia es exclusiva de los mamíferos. [153]
Sistema 5-HT en vertebrados
Los vertebrados son organismos multicelulares del filo Chordata que poseen una columna vertebral y un sistema nervioso . Esto incluye mamíferos, peces, reptiles, aves, etc. En los humanos, el sistema nervioso está compuesto por el sistema nervioso central y periférico , y se sabe poco sobre los mecanismos específicos de los neurotransmisores en la mayoría de los demás vertebrados. Sin embargo, se sabe que, si bien la serotonina está involucrada en el estrés y las respuestas conductuales, también es importante en las funciones cognitivas . [152] La organización cerebral en la mayoría de los vertebrados incluye células 5-HT en el rombencéfalo . [152] Además de esto, la 5-HT se encuentra a menudo en otras secciones del cerebro en vertebrados no placentarios, incluido el prosencéfalo basal y el pretectum . [154] Dado que la ubicación de los receptores de serotonina contribuye a las respuestas conductuales, esto sugiere que la serotonina es parte de vías específicas en vertebrados no placentarios que no están presentes en los organismos amnióticos. [155] Los peces teleósteos y los ratones son los organismos que se utilizan con más frecuencia para estudiar la conexión entre la serotonina y el comportamiento de los vertebrados. Ambos organismos muestran similitudes en el efecto de la serotonina sobre el comportamiento, pero difieren en el mecanismo por el cual se producen las respuestas.
Perros / especies caninas
Existen pocos estudios sobre la serotonina en perros. Un estudio informó que los valores de serotonina eran más altos al amanecer que al anochecer. [156] En otro estudio, los niveles séricos de 5-HT no parecieron estar asociados con la respuesta conductual de los perros a una situación estresante. [157] La relación serotonina/creatinina urinaria en las perras tendió a ser más alta 4 semanas después de la cirugía. Además, la serotonina se correlacionó positivamente con el cortisol y la progesterona, pero no con la testosterona después de la ovariohisterectomía. [158]
Pez teleósteos
Al igual que los vertebrados no placentarios, los peces teleósteos también poseen células 5-HT en otras secciones del cerebro, incluido el prosencéfalo basal . [154] Danio rerio (pez cebra) es una especie de pez teleósteos que se utiliza a menudo para estudiar la serotonina dentro del cerebro. A pesar de que se desconoce mucho sobre los sistemas serotoninérgicos en vertebrados, se conoce su importancia en la moderación del estrés y la interacción social. [159] Se plantea la hipótesis de que AVT y CRF cooperan con la serotonina en el eje hipotálamo-hipofisario-interrenal. [154] Estos neuropéptidos influyen en la plasticidad del teleósteos, afectando su capacidad para cambiar y responder a su entorno. Los peces subordinados en entornos sociales muestran un aumento drástico en las concentraciones de 5-HT. [159] Los niveles altos de 5-HT a largo plazo influyen en la inhibición de la agresión en los peces subordinados. [159]
Ratones
Los investigadores del Departamento de Farmacología y Química Médica utilizaron fármacos serotoninérgicos en ratones macho para estudiar los efectos de determinados fármacos en su comportamiento. [160] Los ratones aislados muestran mayores niveles de comportamiento agonista entre ellos. Los resultados demostraron que los fármacos serotoninérgicos reducen la agresión en ratones aislados al mismo tiempo que aumentan la interacción social. [160] Cada uno de los tratamientos utiliza un mecanismo diferente para atacar la agresión, pero en última instancia todos tienen el mismo resultado. Aunque el estudio muestra que los fármacos serotoninérgicos atacan con éxito a los receptores de serotonina, no muestra detalles de los mecanismos que afectan al comportamiento, ya que todos los tipos de fármacos tendieron a reducir la agresión en ratones machos aislados. [160] Los ratones agresivos que no se encuentran aislados pueden responder de forma diferente a los cambios en la recaptación de serotonina.
Comportamiento
Al igual que en los seres humanos, la serotonina está muy implicada en la regulación del comportamiento en la mayoría de los demás vertebrados. Esto incluye no solo las respuestas y los comportamientos sociales, sino también la influencia en el estado de ánimo. Los defectos en las vías de la serotonina pueden provocar variaciones intensas en el estado de ánimo, así como síntomas de trastornos del estado de ánimo, que pueden estar presentes en más personas que en los seres humanos.
Interacción social
Uno de los aspectos más investigados de la interacción social en el que interviene la serotonina es la agresión. La agresión está regulada por el sistema 5-HT, ya que los niveles de serotonina pueden inducir o inhibir conductas agresivas, como se ha observado en ratones (véase la sección sobre ratones) y cangrejos. [160] Aunque esto es ampliamente aceptado, se desconoce si la serotonina interactúa directa o indirectamente con partes del cerebro que influyen en la agresión y otras conductas. [152] Los estudios de los niveles de serotonina muestran que aumentan y disminuyen drásticamente durante las interacciones sociales, y generalmente se correlacionan con la inhibición o incitación de la conducta agresiva. [161] Se desconoce el mecanismo exacto por el que la serotonina influye en las conductas sociales, ya que las vías en el sistema 5-HT en varios vertebrados pueden diferir en gran medida. [152]
Respuesta a estímulos
La serotonina es importante en las vías de respuesta ambiental, junto con otros neurotransmisores . [162] Específicamente, se ha encontrado que está involucrada en el procesamiento auditivo en entornos sociales, ya que los sistemas sensoriales primarios están conectados a las interacciones sociales. [163] La serotonina se encuentra en la estructura IC del mesencéfalo, que procesa interacciones sociales y vocalizaciones específicas de la especie y no específicas. [163] También recibe proyecciones acústicas que transmiten señales a las regiones de procesamiento auditivo. [163] La investigación ha propuesto que la serotonina da forma a la información auditiva que recibe el IC y, por lo tanto, es influyente en las respuestas a los estímulos auditivos. [163] Esto puede influir en cómo un organismo responde a los sonidos de especies depredadoras u otras especies impactantes en su entorno, ya que la captación de serotonina puede influir en la agresión o la interacción social.
Ánimo
Podemos describir el estado de ánimo no como específico de un estado emocional, sino como asociado con un estado emocional relativamente duradero. La asociación de la serotonina con el estado de ánimo es más conocida por varias formas de depresión y trastornos bipolares en humanos. [153] Los trastornos causados por la actividad serotoninérgica contribuyen potencialmente a los muchos síntomas de la depresión mayor, como el estado de ánimo general, la actividad, los pensamientos suicidas y la disfunción sexual y cognitiva . Los inhibidores selectivos de la recaptación de serotonina (ISRS) son una clase de medicamentos que han demostrado ser un tratamiento eficaz en el trastorno depresivo mayor y son la clase de antidepresivos más recetada. La función de los ISRS es bloquear la recaptación de serotonina, haciendo que haya más serotonina disponible para ser absorbida por la neurona receptora. Los animales han sido estudiados durante décadas para comprender el comportamiento depresivo entre las especies. Uno de los estudios más conocidos, la prueba de natación forzada (FST), se realizó para medir la actividad antidepresiva potencial. [153] Se colocó a las ratas en un recipiente con agua que no se podía escapar, momento en el que se comparó el tiempo que permanecieron inmóviles y el número de conductas activas (como chapotear o trepar) antes y después de administrarles un panel de fármacos antidepresivos. Se demostró que los antidepresivos que inhiben selectivamente la recaptación de NE reducen la inmovilidad y aumentan selectivamente la capacidad de trepar sin afectar la natación. Sin embargo, los resultados de los ISRS también muestran una inmovilidad reducida pero un aumento de la capacidad de nadar sin afectar la capacidad de trepar. Este estudio demostró la importancia de las pruebas de comportamiento para los antidepresivos, ya que pueden detectar fármacos con un efecto sobre el comportamiento central junto con los componentes conductuales de las especies. [153]
Crecimiento y reproducción
En el nematodo C. elegans , la disminución artificial de serotonina o el aumento de octopamina indica un comportamiento típico de un entorno con poca comida: C. elegans se vuelve más activo y se suprimen el apareamiento y la puesta de huevos, mientras que ocurre lo contrario si se aumenta la serotonina o se reduce la octopamina en este animal. [34] La serotonina es necesaria para el comportamiento normal de apareamiento del macho nematodo, [164] y la inclinación a dejar comida para buscar pareja. [165] La señalización serotoninérgica utilizada para adaptar el comportamiento del gusano a los cambios rápidos en el entorno afecta a la señalización similar a la insulina y a la vía de señalización TGF beta , [166] que controlan la adaptación a largo plazo.
En la mosca de la fruta , la insulina regula el azúcar en sangre y actúa como factor de crecimiento . Por lo tanto, en la mosca de la fruta, las neuronas serotoninérgicas regulan el tamaño corporal adulto al afectar la secreción de insulina. [167] [168] La serotonina también se ha identificado como el desencadenante del comportamiento de enjambre en las langostas. [143] En los humanos, aunque la insulina regula el azúcar en sangre y el IGF regula el crecimiento, la serotonina controla la liberación de ambas hormonas, modulando la liberación de insulina de las células beta en el páncreas a través de la serotonina de las proteínas de señalización GTPasa. [44] La exposición a los ISRS durante el embarazo reduce el crecimiento fetal. [169]
Los gusanos C. elegans genéticamente alterados que carecen de serotonina tienen una mayor esperanza de vida reproductiva, pueden volverse obesos y, a veces, presentan un desarrollo detenido en un estado larvario latente . [170] [171]
Envejecimiento y fenotipos relacionados con la edad
Se sabe que la serotonina regula el envejecimiento, el aprendizaje y la memoria. La primera evidencia proviene del estudio de la longevidad en C. elegans . [166] Durante la fase temprana del envejecimiento [ vago ] , el nivel de serotonina aumenta, lo que altera los comportamientos locomotores y la memoria asociativa. [172] El efecto se restaura mediante mutaciones y medicamentos (incluidos mianserina y metiotepina ) que inhiben los receptores de serotonina . La observación no contradice la noción de que el nivel de serotonina disminuye en mamíferos y humanos, lo que generalmente se observa en la fase tardía pero no temprana [ vago ] del envejecimiento.
Mecanismos bioquímicos
Biosíntesis

En los animales y los seres humanos, la serotonina se sintetiza a partir del aminoácido L - triptófano mediante una vía metabólica corta que consta de dos enzimas , la triptófano hidroxilasa (TPH) y la descarboxilasa de aminoácidos aromáticos (DDC), y la coenzima piridoxal fosfato . La reacción mediada por TPH es el paso limitante de la velocidad de la vía.
Se ha demostrado que el TPH existe en dos formas: TPH1 , que se encuentra en varios tejidos , y TPH2 , que es una isoforma específica de las neuronas . [173]
La serotonina se puede sintetizar a partir del triptófano en el laboratorio utilizando Aspergillus niger y Psilocybe coprophila como catalizadores. La primera fase para obtener el 5-hidroxitriptófano requeriría dejar reposar el triptófano en etanol y agua durante 7 días, luego mezclar con suficiente HCl (u otro ácido) para llevar el pH a 3 y luego agregar NaOH para lograr un pH de 13 durante 1 hora. Aspergillus niger sería el catalizador para esta primera fase. La segunda fase para sintetizar el propio triptófano a partir del intermediario 5-hidroxitriptófano requeriría agregar etanol y agua, y dejar reposar durante 30 días esta vez. Los dos pasos siguientes serían los mismos que la primera fase: agregar HCl para lograr un pH de 3 y luego agregar NaOH para lograr un pH muy básico de 13 durante 1 hora. Esta fase utiliza Psilocybe coprophila como catalizador para la reacción. [174]

La serotonina ingerida por vía oral no pasa a las vías serotoninérgicas del sistema nervioso central, porque no cruza la barrera hematoencefálica . [9] Sin embargo, el triptófano y su metabolito 5-hidroxitriptófano (5-HTP), a partir del cual se sintetiza la serotonina, sí cruzan la barrera hematoencefálica. Estos agentes están disponibles como suplementos dietéticos y en diversos alimentos, y pueden ser agentes serotoninérgicos eficaces.
Un producto de la degradación de la serotonina es el ácido 5-hidroxiindolacético (5-HIAA), que se excreta en la orina . En ocasiones, ciertos tumores o cánceres producen serotonina y 5-HIAA en cantidades excesivas , y los niveles de estas sustancias se pueden medir en la orina para detectar estos tumores.
Química analítica
El óxido de indio y estaño se recomienda como material de electrodo en la investigación electroquímica de concentraciones producidas, detectadas o consumidas por microbios . [175] En 1994 se desarrolló una técnica de espectrometría de masas para medir el peso molecular de las serotoninas tanto naturales como sintéticas. [176]
Historia y etimología
Los fisiólogos sabían desde hacía más de un siglo que un material vasoconstrictor aparece en el suero cuando se permite que la sangre coagule. [177] En 1935, el italiano Vittorio Erspamer demostró que un extracto de células enterocromafines hacía que los intestinos se contrajeran. Algunos creían que contenía adrenalina , pero dos años después, Erspamer pudo demostrar que era una amina previamente desconocida , a la que llamó "enteramina". [178] [179] En 1948, Maurice M. Rapport , Arda Green e Irvine Page de la Clínica Cleveland descubrieron una sustancia vasoconstrictora en el suero sanguíneo y, dado que era un agente sérico que afectaba al tono vascular, lo llamaron serotonina. [180]
En 1952, se demostró que la enteramina era la misma sustancia que la serotonina, y como se dilucidó la amplia gama de funciones fisiológicas, la abreviatura 5-HT del nombre químico apropiado 5-hidroxitriptamina se convirtió en el nombre preferido en el campo farmacológico. [181] Los sinónimos de serotonina incluyen: 5-hidroxitriptamina, enteramina, sustancia DS y 3-(β-aminoetil)-5-hidroxiindol. [182] En 1953, Betty Twarog y Page descubrieron la serotonina en el sistema nervioso central. [183] Page consideró que el trabajo de Erspamer en Octopus vulgaris , Discoglossus pictus , Hexaplex trunculus , Bolinus brandaris , Sepia , Mytilus y Ostrea era válido y fundamental para comprender esta sustancia recientemente identificada, pero consideró que sus resultados anteriores en varios modelos, especialmente los de sangre de rata, estaban demasiado confundidos por la presencia de otros productos químicos bioactivos, incluidos algunos otros vasoactivos. [184]
Notas
- ^ Las referencias sobre las funciones de estos receptores están disponibles en las páginas de Wikipedia correspondientes al receptor específico en cuestión.
Referencias
- ^ Calculado utilizando el software Advanced Chemistry Development (ACD/Labs) V11.02 (©1994–2011 ACD/Labs)
- ^ Mazák K, Dóczy V, Kökösi J, Noszál B (abril de 2009). "Especiación de protones y microespeciación de serotonina y 5-hidroxitriptófano". Química y Biodiversidad . 6 (4): 578–590. doi :10.1002/cbdv.200800087. PMID 19353542. S2CID 20543931.
- ^ Pietra S (1958). "[Derivados indólicos. II. Una nueva forma de sintetizar serotonina]". Il Farmaco; Edizione Scientifica (en italiano). 13 (1): 75–79. PMID 13524273.
- ^ Erspamer V (1952). "Ricerche preliminari sulle indolalchilamine e sulle fenilalchilamine degli estratti di pelle di Anfibio". Ricerca Científica . 22 : 694–702.
- ^ Tammisto T (1967). "Aumento de la toxicidad de la 5-hidroxitriptamina por el etanol en ratas y ratones". Annales Medicinae Experimentalis et Biologiae Fenniae . 46 (3): 382–384. PMID 5734241.
- ^ Jones D (2003) [1917]. Roach P, Hartmann J, Setter J (eds.). Diccionario de pronunciación del inglés . Cambridge: Cambridge University Press. ISBN 978-3-12-539683-8.
- ^ "Serotonina". Dictionary.com Unabridged (en línea). nd
- ^ "Serotonina". Diccionario Merriam-Webster.com . Merriam-Webster.
- ^ ab Young SN (noviembre de 2007). "Cómo aumentar la serotonina en el cerebro humano sin fármacos". Journal of Psychiatry & Neuroscience . 32 (6): 394–399. PMC 2077351 . PMID 18043762.
- ^ "Los microbios ayudan a producir serotonina en el intestino". Instituto Tecnológico de California . 9 de abril de 2015. Consultado el 3 de junio de 2022 .
- ^ King MW. "Serotonina". Página de bioquímica médica . Facultad de Medicina de la Universidad de Indiana . Consultado el 1 de diciembre de 2009 .
- ^ Berger M, Gray JA, Roth BL (2009). "La biología expandida de la serotonina". Revista Anual de Medicina . 60 : 355–366. doi :10.1146/annurev.med.60.042307.110802. PMC 5864293 . PMID 19630576.
- ^ Schlienger RG, Meier CR (2003). "Efecto de los inhibidores selectivos de la recaptación de serotonina en la activación plaquetaria: ¿pueden prevenir el infarto agudo de miocardio?". American Journal of Cardiovascular Drugs . 3 (3): 149–162. doi :10.2165/00129784-200303030-00001. PMID 14727927. S2CID 23986530.
- ^ Kling A (2013). 5-HT2A: un receptor de serotonina con un posible papel en las enfermedades articulares (PDF) (Tesis). Universidad de Umeå. ISBN 978-91-7459-549-9.
- ^ Yano JM, Yu K, Donaldson GP, Shastri GG, Ann P, Ma L, et al. (abril de 2015). "Las bacterias autóctonas de la microbiota intestinal regulan la biosíntesis de serotonina del huésped". Cell . 161 (2): 264–276. doi :10.1016/j.cell.2015.02.047. PMC 4393509 . PMID 25860609.
- ^ Sangare A, Dubourget R, Geoffroy H, Gallopin T, Rancillac A (octubre de 2016). "La serotonina modula de manera diferencial las entradas sinápticas excitatorias e inhibidoras a las neuronas promotoras del sueño putativas del núcleo preóptico ventrolateral" (PDF) . Neurofarmacología . 109 : 29–40. doi :10.1016/j.neuropharm.2016.05.015. PMID 27238836.
- ^ Rancillac A (noviembre de 2016). "Serotonina y neuronas promotoras del sueño". Oncotarget . 7 (48): 78222–78223. doi :10.18632/oncotarget.13419. PMC 5346632 . PMID 27861160.
- ^ Vanhoutte PM (febrero de 1987). "Serotonina y la pared vascular". Revista internacional de cardiología . 14 (2): 189–203. doi :10.1016/0167-5273(87)90008-8. PMID 3818135.
- ^ ab Huser A, Rohwedder A, Apostolopoulou AA, Widmann A, Pfitzenmaier JE, Maiolo EM, et al. (2012). Zars T (ed.). "El sistema nervioso central serotoninérgico de la larva de Drosophila: anatomía y función conductual". PLOS ONE . 7 (10): e47518. Bibcode :2012PLoSO...747518H. doi : 10.1371/journal.pone.0047518 . PMC 3474743 . PMID 23082175.
- ^ abc Ramakrishna A, Giridhar P, Ravishankar GA (junio de 2011). "Fitoserotonina: una revisión". Plant Signaling & Behavior . 6 (6): 800–809. Bibcode :2011PlSiB...6..800A. doi :10.4161/psb.6.6.15242. PMC 3218476 . PMID 21617371.
- ^ ab Chen J, Lariviere WR (octubre de 2010). "Los efectos nociceptivos y antinociceptivos de la inyección y la terapia con veneno de abeja: un arma de doble filo". Progress in Neurobiology . 92 (2): 151–183. doi :10.1016/j.pneurobio.2010.06.006. PMC 2946189 . PMID 20558236.
- ^ abcde Erspamer V (1966). "Presencia de indolalquilaminas en la naturaleza". 5-Hidroxitriptamina y indolalquilaminas relacionadas . Berlín , Heidelberg : Springer Berlin Heidelberg . págs. 132–181. doi :10.1007/978-3-642-85467-5_4. ISBN 978-3-642-85469-9.
- ^ ab McGowan K, Kane A, Asarkof N, Wicks J, Guerina V, Kellum J, et al. (agosto de 1983). "Entamoeba histolytica provoca secreción intestinal: papel de la serotonina". Science . 221 (4612): 762–764. Bibcode :1983Sci...221..762M. doi :10.1126/science.6308760. PMID 6308760.
- ^ ab Feldman JM, Lee EM (octubre de 1985). "Contenido de serotonina en los alimentos: efecto sobre la excreción urinaria de ácido 5-hidroxiindolacético". The American Journal of Clinical Nutrition . 42 (4): 639–643. doi : 10.1093/ajcn/42.4.639 . PMID 2413754.
- ^ González-Flores D, Velardo B, Garrido M, et al. (2011). "La ingestión de ciruelas japonesas (Prunus salicina Lindl. cv. Crimson Globe) aumenta los niveles urinarios de 6-sulfatoximelatonina y de capacidad antioxidante total en humanos jóvenes, de mediana edad y ancianos: Caracterización nutricional y funcional de su contenido". Revista de Investigación en Alimentación y Nutrición . 50 (4): 229–236.
- ^ Rychkov DA, Hunter S, Kovalskii VY, Lomzov AA, Pulham CR, Boldyreva EV (julio de 2016). "Hacia una comprensión de la cristalización a partir de una solución. Estudios DFT de cristales de serotonina multicomponente". Química computacional y teórica . 1088 : 52–61. doi :10.1016/j.comptc.2016.04.027.
- ^ Naeem M, Chadeayne AR, Golen JA, Manke DR (abril de 2022). "Estructura cristalina de la serotonina". Acta Crystallographica Sección E . 78 (Pt 4): 365–368. Código Bibliográfico :2022AcCrE..78..365N. doi :10.1107/S2056989022002559. PMC 8983975 . PMID 35492269.
- ^ Rychkov D, Boldyreva EV, Tumanov NA (septiembre de 2013). "Una nueva estructura de una sal de serotonina: comparación y análisis conformacional de todos los complejos de serotonina conocidos". Acta Crystallographica Sección C . 69 (Pt 9): 1055–1061. Bibcode :2013AcCrC..69.1055R. doi :10.1107/S0108270113019823. PMID 24005521.
- ^ Mohammad-Zadeh LF, Moses L, Gwaltney-Brant SM (junio de 2008). "Serotonina: una revisión". Revista de farmacología y terapéutica veterinaria . 31 (3): 187–199. doi : 10.1111/j.1365-2885.2008.00944.x . PMID 18471139.
- ^ Vaseghi S, Arjmandi-Rad S, Eskandari M, Ebrahimnejad M, Kholghi G, Zarrindast MR (febrero de 2022). "Modulación del papel de la señalización serotoninérgica en el sueño y la memoria". Pharmacological Reports . 74 (1): 1–26. doi :10.1007/s43440-021-00339-8. PMID 34743316.
- ^ ab Kravitz EA (septiembre de 1988). "Control hormonal del comportamiento: aminas y sesgo del comportamiento en langostas". Science . 241 (4874): 1775–1781. Bibcode :1988Sci...241.1775K. doi :10.1126/science.2902685. PMID 2902685.
- ^ Sinclair-Wilson A, Lawrence A, Ferezou I, Cartonnet H, Mailhes C, Garel S, et al. (agosto de 2023). "La plasticidad de los axones talamocorticales está regulada por los niveles de serotonina modulados por el parto prematuro". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 120 (33): e2301644120. Bibcode :2023PNAS..12001644S. doi :10.1073/pnas.2301644120. PMC 10438379 . PMID 37549297.
- ^ ab Zifa E, Fillion G (septiembre de 1992). "Receptores de 5-hidroxitriptamina". Revisiones farmacológicas . 44 (3): 401–458. PMID 1359584.
- ^ ab Srinivasan S, Sadegh L, Elle IC, Christensen AG, Faergeman NJ, Ashrafi K (junio de 2008). "La serotonina regula la grasa y la alimentación de C. elegans a través de mecanismos moleculares independientes". Metabolismo celular . 7 (6): 533–544. doi :10.1016/j.cmet.2008.04.012. PMC 2495008 . PMID 18522834.
- ^ Ramakrishna A, Ravishankar GA (noviembre de 2011). "Influencia de las señales de estrés abiótico en los metabolitos secundarios de las plantas". Plant Signaling & Behavior . 6 (11). Informa : 1720–1731. Bibcode :2011PlSiB...6.1720A. doi : 10.4161/psb.6.11.17613 . PMC 3329344 . PMID 22041989.
- ^ Whitaker R, Cosgrove L (2015). Psiquiatría bajo la influencia: corrupción institucional, daño social y prescripciones para la reforma. Springer. págs. 55-56. ISBN 978-1-137-51602-2.
- ^ Moncrieff J, Cooper RE, Stockmann T, Amendola S, Hengartner MP, Horowitz MA (agosto de 2023). "La teoría de la serotonina de la depresión: una revisión sistemática general de la evidencia". Psiquiatría molecular . 28 (8). Nature Publishing Group: 3243–3256. doi : 10.1038/s41380-022-01661-0 . PMC 10618090 . PMID 35854107. S2CID 250646781.
- ^ Ghaemi N (2022), ¿Se ha desacreditado la hipótesis de la serotonina? , consultado el 2 de mayo de 2023
- ^ Bortolato M, Chen K, Shih JC (2010). "La degradación de la serotonina: papel de la MAO". Handbook of Behavioral Neuroscience . Vol. 21. Elsevier. págs. 203–218. doi :10.1016/s1569-7339(10)70079-5. ISBN 978-0-12-374634-4.
- ^ Matthes S, Mosienko V, Bashammakh S, Alenina N, Bader M (2010). "La triptófano hidroxilasa como nueva diana para el tratamiento de los trastornos depresivos". Farmacología . 85 (2): 95–109. doi :10.1159/000279322. PMID 20130443.
- ^ Prah A, Purg M, Stare J, Vianello R, Mavri J (septiembre de 2020). "Cómo la monoaminooxidasa A descompone la serotonina: una simulación empírica del enlace de valencia del paso reactivo". The Journal of Physical Chemistry B . 124 (38): 8259–8265. doi :10.1021/acs.jpcb.0c06502. PMC 7520887 . PMID 32845149.
- ^ Hannon J, Hoyer D (diciembre de 2008). "Biología molecular de los receptores 5-HT". Behavioural Brain Research . 195 (1): 198–213. doi :10.1016/j.bbr.2008.03.020. PMID 18571247. S2CID 46043982.
- ^ ab Zhou M, Engel K, Wang J (enero de 2007). "Evidencia de una contribución significativa de un transportador de monoaminas recientemente identificado (PMAT) a la captación de serotonina en el cerebro humano". Farmacología bioquímica . 73 (1): 147–154. doi :10.1016/j.bcp.2006.09.008. PMC 1828907 . PMID 17046718.
- ^ abc Paulmann N, Grohmann M, Voigt JP, Bert B, Vowinckel J, Bader M, et al. (octubre de 2009). O'Rahilly S (ed.). "La serotonina intracelular modula la secreción de insulina de las células beta pancreáticas mediante serotonina proteica". PLOS Biology . 7 (10): e1000229. doi : 10.1371/journal.pbio.1000229 . PMC 2760755 . PMID 19859528.
- ^ Walther DJ, Peter JU, Winter S, Höltje M, Paulmann N, Grohmann M, et al. (diciembre de 2003). "La serotonina de pequeñas GTPasas es una vía de transducción de señales que desencadena la liberación de gránulos alfa de plaquetas". Cell . 115 (7): 851–862. doi : 10.1016/S0092-8674(03)01014-6 . PMID 14697203. S2CID 16847296.
- ^ Watts SW, Priestley JR, Thompson JM (mayo de 2009). "Serotonilación de proteínas vasculares importantes para la contracción". PLOS ONE . 4 (5): e5682. Bibcode :2009PLoSO...4.5682W. doi : 10.1371/journal.pone.0005682 . PMC 2682564 . PMID 19479059.
- ^ Roth BL, Driscol J (12 de enero de 2011). "PDSP Ki Database". Programa de detección de drogas psicoactivas (PDSP) . Universidad de Carolina del Norte en Chapel Hill y el Instituto Nacional de Salud Mental de los Estados Unidos. Archivado desde el original el 8 de noviembre de 2013. Consultado el 17 de diciembre de 2013 .
- ^ Bortolozzi A, Díaz-Mataix L, Scorza MC, Celada P, Artigas F (diciembre de 2005). "La activación de los receptores 5-HT en la corteza prefrontal potencia la actividad dopaminérgica". Revista de neuroquímica . 95 (6): 1597-1607. doi :10.1111/j.1471-4159.2005.03485.x. hdl : 10261/33026 . PMID 16277612. S2CID 18350703.
- ^ Moro C, Edwards L, Chess-Williams R (noviembre de 2016). "Mejora de la actividad contráctil del urotelio y la lámina propia porcinos a través del receptor 5-HT2A". Revista Internacional de Urología . 23 (11): 946–951. doi : 10.1111/iju.13172 . PMID 27531585.
- ^ "Neuron de Economo – NeuronBank". neuronbank.org .[¿ Fuente médica poco confiable? ]
- ^ Millan MJ, Gobert A, Lejeune F, Dekeyne A, Newman-Tancredi A, Pasteau V, et al. (septiembre de 2003). "El nuevo agonista de la melatonina agomelatina (S20098) es un antagonista de los receptores 5-hidroxitriptamina2C, cuyo bloqueo mejora la actividad de las vías dopaminérgicas y adrenérgicas frontocorticales". The Journal of Pharmacology and Experimental Therapeutics . 306 (3): 954–964. doi :10.1124/jpet.103.051797. PMID 12750432. S2CID 18753440.
- ^ Gonzalez R, Chávez-Pascacio K, Meneses A (septiembre de 2013). "Role of 5-HT5A receptors in the consolidation of memory" (Papel de los receptores 5-HT5A en la consolidación de la memoria). Behavioural Brain Research . 252 : 246–251. doi :10.1016/j.bbr.2013.05.051. PMID 23735322. S2CID 140204585.
- ^ Nautiyal KM, Hen R (2017). "Receptores de serotonina en la depresión: de A a B". F1000Research . 6 : 123. doi : 10.12688/f1000research.9736.1 . PMC 5302148 . PMID 28232871.
- ^ Frazer A, Hensler JG (1999). "La comprensión de la organización neuroanatómica de las células serotoninérgicas en el cerebro proporciona información sobre las funciones de este neurotransmisor". En Siegel GJ, Agranoff, Bernard W, Fisher SK, Albers RW, Uhler MD (eds.). Neuroquímica básica (sexta edición). Lippincott Williams & Wilkins. ISBN 978-0-397-51820-3
En 1964, Dahlstrom y Fuxe (discutidos en [2]), utilizando la
técnica de histofluorescencia de Falck-Hillarp
, observaron que la mayoría del soma serotoninérgicose encuentra en grupos de cuerpos celulares, que previamente habían sido designados como núcleos del rafe.
- ^ Binder MD, Hirokawa N (2009). Enciclopedia de neurociencia . Berlín: Springer. Pág. 705. ISBN. 978-3-540-23735-8.
- ^ El grupo de neuronas de los núcleos del rafe se encuentra a lo largo del tronco encefálico desde las etiquetas ' Mesoencéfalo ' hasta ' Oblongada ', centrado en el puente . ( Ver imagen relevante ).
- ^ Müller CP, Jacobs BL, eds. (2009). Manual de la neurobiología conductual de la serotonina (1.ª ed.). Londres: Academic. págs. 51–59. ISBN 978-0-12-374634-4.
- ^ Passos AD, Herculano AM, Oliveira KR, de Lima SM, Rocha FA, Freitas HR, et al. (octubre de 2019). "Regulación del sistema serotoninérgico por kainato en la retina aviar". Neurobiología Celular y Molecular . 39 (7): 1039–1049. doi :10.1007/s10571-019-00701-8. PMID 31197744. S2CID 189763144.
- ^ Sommer C (2009). "Serotonina en el dolor y su control". En Müller CP, Jacobs BL (eds.). Manual de la neurobiología conductual de la serotonina (1.ª ed.). Londres: Academic. págs. 457–460. ISBN 978-0-12-374634-4.
- ^ Hensler JG (2009). "Serotonina en el modo y las emociones". En Müller CP, Jacobs BL (eds.). Manual de la neurobiología conductual de la serotonina (1.ª ed.). Londres: Academic. págs. 367–399. ISBN 978-0-12-374634-4.
- ^ Andrews PW, Bharwani A, Lee KR, Fox M, Thomson JA (abril de 2015). "¿La serotonina es un estimulante o un depresor? La evolución del sistema serotoninérgico y su papel en la depresión y la respuesta antidepresiva". Neuroscience and Biobehavioral Reviews . 51 : 164–188. doi :10.1016/j.neubiorev.2015.01.018. PMID 25625874. S2CID 23980182.
- ^ Stahl SM, Mignon L, Meyer JM (marzo de 2009). "¿Qué viene primero: el tratamiento antipsicótico atípico o el riesgo cardiometabólico?". Acta Psychiatrica Scandinavica . 119 (3): 171–179. doi : 10.1111/j.1600-0447.2008.01334.x . PMID 19178394. S2CID 24035040.
- ^ Buckland PR, Hoogendoorn B, Guy CA, Smith SK, Coleman SL, O'Donovan MC (marzo de 2005). "Baja expresión génica conferida por asociación de un alelo del gen del receptor 5-HT2C con aumento de peso inducido por antipsicóticos". The American Journal of Psychiatry . 162 (3): 613–615. doi :10.1176/appi.ajp.162.3.613. PMID 15741483.
- ^ Holmes MC, French KL, Seckl JR (junio de 1997). "Desregulación de los ritmos diurnos de la serotonina 5-HT2C y la expresión génica del receptor de corticosteroides en el hipocampo con restricción de alimentos y glucocorticoides". The Journal of Neuroscience . 17 (11): 4056–4065. doi :10.1523/JNEUROSCI.17-11-04056.1997. PMC 6573558 . PMID 9151722.
- ^ Leibowitz SF (1990). "El papel de la serotonina en los trastornos alimentarios". Drugs . 39 (Supl 3): 33–48. doi :10.2165/00003495-199000393-00005. PMID 2197074. S2CID 8612545.
- ^ McGuire, Michael (2013) "Creer, la neurociencia de las fantasías, los miedos y las contradicciones" (Prometius Books)
- ^ Caspi N, Modai I, Barak P, Waisbourd A, Zbarsky H, Hirschmann S, et al. (marzo de 2001). "Aumento con pindolol en pacientes esquizofrénicos agresivos: un estudio aleatorizado, cruzado y doble ciego". Psicofarmacología Clínica Internacional . 16 (2): 111–115. doi :10.1097/00004850-200103000-00006. PMID 11236069. S2CID 24822810.
- ^ Ito Z, Aizawa I, Takeuchi M, Tabe M, Nakamura T (diciembre de 1975). "[Actas: Estudio de la motilidad gastrointestinal utilizando un transductor de fuerza extraluminal. 6. Observación de la motilidad gástrica y duodenal utilizando motilina sintética]". Nihon Heikatsukin Gakkai Zasshi . 11 (4): 244–246. PMID 1232434.
- ^ Lesch KP, Bengel D, Heils A, Sabol SZ, Greenberg BD, Petri S, et al. (noviembre de 1996). "Asociación de rasgos relacionados con la ansiedad con un polimorfismo en la región reguladora del gen transportador de serotonina". Science . 274 (5292): 1527–1531. Bibcode :1996Sci...274.1527L. doi :10.1126/science.274.5292.1527. PMID 8929413. S2CID 35503987.
- ^ Beattie DT, Smith JA (mayo de 2008). "Farmacología de la serotonina en el tracto gastrointestinal: una revisión". Archivos de farmacología de Naunyn-Schmiedeberg . 377 (3): 181–203. doi :10.1007/s00210-008-0276-9. PMID 18398601. S2CID 32820765.
- ^ Rang HP (2003). Farmacología . Edimburgo: Churchill Livingstone. pág. 187. ISBN. 978-0-443-07145-4.
- ^ de Wit R, Aapro M, Blower PR (septiembre de 2005). "¿Existe una base farmacológica para las diferencias en la eficacia de los antagonistas del receptor 5-HT3 en pacientes refractarios?". Quimioterapia y farmacología del cáncer . 56 (3): 231–238. doi :10.1007/s00280-005-1033-0. PMID 15838653. S2CID 27576150.
- ^ ab Lauweryns JM, Cokelaere J, Theunynck P (abril de 1973). "Cuerpos neuroepiteliales productores de serotonina en la mucosa respiratoria del conejo". Science . 180 (4084): 410–413. Bibcode :1973Sci...180..410L. doi :10.1126/science.180.4084.410. PMID 4121716. S2CID 2809307.
- ^ Pastor LM, Ballesta J, Perez-Tomas R, Marin JA, Hernandez F, Madrid JF (junio de 1987). "Localización inmunocitoquímica de la serotonina en el pulmón de reptiles". Cell and Tissue Research . 248 (3). Springer Science and Business Media LLC: 713–715. doi :10.1007/bf00216504. PMID 3301000. S2CID 9871728.
- ^ ab Sonstegard KS, Mailman RB, Cheek JM, Tomlin TE, DiAugustine RP (noviembre de 1982). "Caracterización morfológica y citoquímica de los cuerpos neuroepiteliales en el pulmón fetal de conejo. I. Estudios de cuerpos neuroepiteliales aislados". Experimental Lung Research . 3 (3–4): 349–377. doi :10.3109/01902148209069663. PMID 6132813.
- ^ Chang W, Kanda H, Ikeda R, Ling J, DeBerry JJ, Gu JG (septiembre de 2016). "El disco de Merkel es una sinapsis serotoninérgica en la epidermis para transmitir señales táctiles en mamíferos". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 113 (37): E5491–E5500. Bibcode :2016PNAS..113E5491C. doi : 10.1073/pnas.1610176113 . PMC 5027443 . PMID 27573850.
- ^ Frost M, Andersen TE, Yadav V, Brixen K, Karsenty G, Kassem M (marzo de 2010). "Los pacientes con fenotipo de masa ósea elevada debido a la mutación Lrp5-T253I tienen niveles plasmáticos bajos de serotonina". Journal of Bone and Mineral Research . 25 (3): 673–675. doi : 10.1002/jbmr.44 . PMID 20200960. S2CID 24280062.
- ^ Rosen CJ (febrero de 2009). "Adentrándonos en la biología ósea: los secretos de la serotonina". Nature Medicine . 15 (2): 145–146. doi :10.1038/nm0209-145. PMID 19197289. S2CID 5489589.
- ^ Mödder UI, Achenbach SJ, Amin S, Riggs BL, Melton LJ, Khosla S (febrero de 2010). "Relación de los niveles séricos de serotonina con la densidad ósea y los parámetros estructurales en mujeres". Journal of Bone and Mineral Research . 25 (2): 415–422. doi :10.1359/jbmr.090721. PMC 3153390 . PMID 19594297.
- ^ Frost M, Andersen T, Gossiel F, Hansen S, Bollerslev J, van Hul W, et al. (agosto de 2011). "Niveles de serotonina, esclerostina, marcadores de recambio óseo, así como densidad ósea y microarquitectura en pacientes con fenotipo de masa ósea alta debido a una mutación en Lrp5". Journal of Bone and Mineral Research . 26 (8): 1721–1728. doi : 10.1002/jbmr.376 . PMID 21351148.
- ^ Kode A, Mosialou I, Silva BC, Rached MT, Zhou B, Wang J, et al. (octubre de 2012). "FOXO1 orquesta la función de supresión ósea de la serotonina derivada del intestino". The Journal of Clinical Investigation . 122 (10): 3490–3503. doi :10.1172/JCI64906. PMC 3461930 . PMID 22945629.
- ^ Yadav VK, Balaji S, Suresh PS, Liu XS, Lu X, Li Z, et al. (marzo de 2010). "La inhibición farmacológica de la síntesis de serotonina derivada del intestino es un posible tratamiento anabólico óseo para la osteoporosis". Nature Medicine . 16 (3): 308–312. doi :10.1038/nm.2098. PMC 2836724 . PMID 20139991.
- ^ Sugisawa E, Takayama Y, Takemura N, Kondo T, Hatakeyama S, Kumagai Y, et al. (agosto de 2020). "La detección de ARN mediante Gut Piezo1 es esencial para la síntesis de serotonina sistémica". Celúla . 182 (3): 609–624.e21. doi : 10.1016/j.cell.2020.06.022 . PMID 32640190.
- ^ Ozanne SE, Hales CN (enero de 2004). "Longitud vital: crecimiento compensatorio y obesidad en ratones macho". Nature . 427 (6973): 411–412. Bibcode :2004Natur.427..411O. doi :10.1038/427411b. PMID 14749819. S2CID 40256021.
- ^ Lewis DS, Bertrand HA, McMahan CA, McGill HC, Carey KD, Masoro EJ (octubre de 1986). "La ingesta de alimentos antes del destete influye en la adiposidad de los babuinos adultos jóvenes". The Journal of Clinical Investigation . 78 (4): 899–905. doi :10.1172/JCI112678. PMC 423712 . PMID 3760191.
- ^ Hahn P (julio de 1984). "Efecto del tamaño de la camada sobre el colesterol plasmático y la insulina y algunas enzimas del hígado y del tejido adiposo en roedores adultos". The Journal of Nutrition . 114 (7): 1231–1234. doi :10.1093/jn/114.7.1231. PMID 6376732.
- ^ Popa D, Léna C, Alexandre C, Adrien J (abril de 2008). "Síndrome duradero de depresión producido por la reducción de la captación de serotonina durante el desarrollo posnatal: evidencia del sueño, el estrés y el comportamiento". The Journal of Neuroscience . 28 (14): 3546–3554. doi :10.1523/JNEUROSCI.4006-07.2008. PMC 6671102 . PMID 18385313.
- ^ Maciag D, Simpson KL, Coppinger D, Lu Y, Wang Y, Lin RC, et al. (enero de 2006). "La exposición neonatal a antidepresivos tiene efectos duraderos en el comportamiento y los circuitos de serotonina". Neuropsicofarmacología . 31 (1): 47–57. doi :10.1038/sj.npp.1300823. PMC 3118509 . PMID 16012532.
- ^ Maciag D, Williams L, Coppinger D, Paul IA (febrero de 2006). "La exposición neonatal al citalopram produce cambios duraderos en el comportamiento que se revierten con el tratamiento con imipramina en adultos". Revista Europea de Farmacología . 532 (3): 265–269. doi :10.1016/j.ejphar.2005.12.081. PMC 2921633 . PMID 16483567.
- ^ Holden C (octubre de 2004). "Neurociencia. El tratamiento con Prozac de ratones recién nacidos aumenta la ansiedad". Science . 306 (5697): 792. doi : 10.1126/science.306.5697.792 . PMID 15514122.
- ^ Ansorge MS, Zhou M, Lira A, Hen R, Gingrich JA (octubre de 2004). "El bloqueo del transportador 5-HT en etapas tempranas de la vida altera el comportamiento emocional en ratones adultos". Science . 306 (5697): 879–881. Bibcode :2004Sci...306..879A. doi : 10.1126/science.1101678 . PMID 15514160.
- ^ Lesurtel M, Graf R, Aleil B, Walther DJ, Tian Y, Jochum W, et al. (abril de 2006). "La serotonina derivada de plaquetas media la regeneración del hígado". Science . 312 (5770): 104–107. Bibcode :2006Sci...312..104L. doi :10.1126/science.1123842. PMID 16601191. S2CID 43189753.
- ^ Matondo RB, Punt C, Homberg J, Toussaint MJ, Kisjes R, Korporaal SJ, et al. (abril de 2009). "La eliminación del transportador de serotonina en ratas altera la homeostasis de la serotonina sin perjudicar la regeneración hepática". American Journal of Physiology. Fisiología gastrointestinal y hepática . 296 (4): G963–G968. doi :10.1152/ajpgi.90709.2008. PMID 19246633. Archivado desde el original el 28 de diciembre de 2019. Consultado el 5 de diciembre de 2019 .
- ^ Collet C, Schiltz C, Geoffroy V, Maroteaux L, Launay JM, de Vernejoul MC (febrero de 2008). "El receptor de serotonina 5-HT2B controla la masa ósea a través del reclutamiento y la proliferación de osteoblastos". FASEB Journal . 22 (2): 418–427. doi : 10.1096/fj.07-9209com . PMC 5409955 . PMID 17846081.
- ^ Yadav VK, Ryu JH, Suda N, Tanaka KF, Gingrich JA, Schütz G, et al. (noviembre de 2008). "Lrp5 controla la formación ósea inhibiendo la síntesis de serotonina en el duodeno". Cell . 135 (5): 825–837. doi :10.1016/j.cell.2008.09.059. PMC 2614332 . PMID 19041748.
- "Se necesitan agallas para formar huesos, descubren los científicos". ScienceDaily (nota de prensa). 1 de diciembre de 2008.
- ^ McDuffie JE, Motley ED, Limbird LE, Maleque MA (marzo de 2000). "La 5-hidroxitriptamina estimula la fosforilación de la activación de la proteína quinasa activada por mitógeno p44/p42 en cultivos de células endoteliales aórticas bovinas". Journal of Cardiovascular Pharmacology . 35 (3): 398–402. doi : 10.1097/00005344-200003000-00008 . PMID 10710124.
- ^ Noguchi M, Furukawa KT, Morimoto M (diciembre de 2020). "Células neuroendocrinas pulmonares: fisiología, homeostasis tisular y enfermedad". Modelos y mecanismos de enfermedades . 13 (12): dmm046920. doi :10.1242/dmm.046920. PMC 7774893 . PMID 33355253.
- ^ Fuller RW (1980). "Farmacología de las neuronas serotoninérgicas centrales". Revisión anual de farmacología y toxicología . 20 : 111–127. doi :10.1146/annurev.pa.20.040180.000551. PMID 6992697.
- ^ Titeler M, Lyon RA, Glennon RA (1988). "La evidencia de unión de radioligandos implica al receptor 5-HT2 del cerebro como un sitio de acción para el LSD y los alucinógenos de fenilisopropilamina". Psicofarmacología . 94 (2): 213–216. doi :10.1007/BF00176847. PMID 3127847. S2CID 24179554.
- ^ Nichols DE (2000). "El papel de las neuronas serotoninérgicas y los receptores 5-HT en la acción de los alucinógenos". En Baumgarten HG, Gothert M (eds.). Neuronas serotoninérgicas y receptores 5-HT en el sistema nervioso central . Santa Clara, CA: Springer-Verlag TELOS. ISBN 978-3-540-66715-5.
- ^ Kapur S, Seeman P (2002). "Los antagonistas del receptor NMDA, la ketamina y el PCP, tienen efectos directos sobre los receptores de dopamina D(2) y serotonina 5-HT(2): implicaciones para los modelos de esquizofrenia". Molecular Psychiatry . 7 (8): 837–844. doi : 10.1038/sj.mp.4001093 . PMID 12232776.
- ^ Johnson MP, Hoffman AJ, Nichols DE (diciembre de 1986). "Efectos de los enantiómeros de MDA, MDMA y análogos relacionados en la liberación de [3H]serotonina y [3H]dopamina a partir de cortes de cerebro de rata superfusionados". Revista Europea de Farmacología . 132 (2–3): 269–276. doi :10.1016/0014-2999(86)90615-1. PMID 2880735.
- ^ Goodman LS, Brunton LL, Chabner B, Knollmann BC (2001). Bases farmacológicas de la terapéutica de Goodman y Gilman . Nueva York: McGraw-Hill. págs. 459–461. ISBN. 978-0-07-162442-8.
- ^ Benmansour S, Cecchi M, Morilak DA, Gerhardt GA, Javors MA, Gould GG, et al. (diciembre de 1999). "Efectos de los tratamientos antidepresivos crónicos en la función, densidad y nivel de ARNm del transportador de serotonina". The Journal of Neuroscience . 19 (23): 10494–10501. doi :10.1523/JNEUROSCI.19-23-10494.1999. PMC 6782424 . PMID 10575045.
- ^ Beitchman JH, Baldassarra L, Mik H, De Luca V, King N, Bender D, et al. (junio de 2006). "Polimorfismos del transportador de serotonina y agresión infantil persistente y generalizada". The American Journal of Psychiatry . 163 (6): 1103–1105. doi :10.1176/appi.ajp.163.6.1103. PMID 16741214.
- ^ Pezawas L, Meyer-Lindenberg A, Drabant EM, Verchinski BA, Munoz KE, Kolachana BS, et al. (junio de 2005). "El polimorfismo 5-HTTLPR afecta las interacciones entre la amígdala y el cíngulo humano: un mecanismo de susceptibilidad genética a la depresión". Nature Neuroscience . 8 (6): 828–834. doi :10.1038/nn1463. PMID 15880108. S2CID 1864631.
- ^ Schinka JA, Busch RM, Robichaux-Keene N (febrero de 2004). "Un metaanálisis de la asociación entre el polimorfismo del gen transportador de serotonina (5-HTTLPR) y la ansiedad como rasgo". Psiquiatría molecular . 9 (2): 197–202. doi : 10.1038/sj.mp.4001405 . PMID 14966478.
- ^ ab New AM, Nelson S, Leung JG (1 de octubre de 2015). Alexander E, Susla GM (eds.). "Emergencias psiquiátricas en la unidad de cuidados intensivos". AACN Advanced Critical Care . 26 (4): 285–293, cuestionario 294–295. doi :10.4037/NCI.0000000000000104. PMID 26484986.
- ^ Isbister GK, Bowe SJ, Dawson A, Whyte IM (2004). "Toxicidad relativa de los inhibidores selectivos de la recaptación de serotonina (ISRS) en caso de sobredosis". Revista de toxicología. Toxicología clínica . 42 (3): 277–285. doi :10.1081/CLT-120037428. PMID 15362595. S2CID 43121327.
- ^ Dunkley EJ, Isbister GK, Sibbritt D, Dawson AH, Whyte IM (septiembre de 2003). "Los criterios de toxicidad de la serotonina de Hunter: reglas de decisión diagnóstica simples y precisas para la toxicidad de la serotonina". QJM . 96 (9): 635–642. doi : 10.1093/qjmed/hcg109 . PMID 12925718.
- ^ Frank C (julio de 2008). "Reconocimiento y tratamiento del síndrome serotoninérgico". Canadian Family Physician . 54 (7): 988–992. PMC 2464814 . PMID 18625822.
- ^ Boyer EW, Shannon M (marzo de 2005). "El síndrome de la serotonina". The New England Journal of Medicine . 352 (11): 1112–1120. doi :10.1056/NEJMra041867. PMID 15784664.
- ^ ab Baskin SI (1991). Principios de toxicología cardíaca. Boca Raton: CRC Press. ISBN 978-0-8493-8809-5. Recuperado el 3 de febrero de 2010 .
- ^ Jähnichen S, Horowski R, Pertz H. "La pergolida y la cabergolina, pero no la lisurida, muestran eficacia agonista en los receptores de serotonina 5-HT2B" (PDF) . Consultado el 3 de febrero de 2010 .
- ^ Comité Asesor sobre Reacciones Adversas a Medicamentos, Australia (2004). "Valvulopatía cardíaca con pergolida". Boletín Aust sobre Reacciones Adversas a Medicamentos 23 (4). Archivado desde el original el 27 de junio de 2012.
- ^ Schade R, Andersohn F, Suissa S, Haverkamp W, Garbe E (enero de 2007). "Agonistas de la dopamina y riesgo de insuficiencia valvular cardíaca". The New England Journal of Medicine . 356 (1): 29–38. doi : 10.1056/NEJMoa062222 . PMID 17202453.
- ^ Zanettini R, Antonini A, Gatto G, Gentile R, Tesei S, Pezzoli G (enero de 2007). "Valvulopatías y uso de agonistas de la dopamina para la enfermedad de Parkinson". The New England Journal of Medicine . 356 (1): 39–46. doi : 10.1056/NEJMoa054830 . PMID 17202454.
- ^ "Aviso de salud pública de la Administración de Alimentos y Medicamentos". Administración de Alimentos y Medicamentos . 29 de marzo de 2007 . Consultado el 7 de febrero de 2010 .
- ^ "MedWatch – Alertas de información de seguridad de 2007. Permax (pergolida) y equivalentes genéricos". Administración de Alimentos y Medicamentos de los Estados Unidos . 29 de marzo de 2007. Consultado el 30 de marzo de 2007 .
- ^ Tyler VE (septiembre de 1958). "Presencia de serotonina en un hongo alucinógeno". Science . 128 (3326): 718. Bibcode :1958Sci...128..718T. doi :10.1126/science.128.3326.718. PMID 13580242.
- ^ Johnson DJ, Sanderson H, Brain RA, Wilson CJ, Solomon KR (mayo de 2007). "Toxicidad y peligro de los antidepresivos inhibidores selectivos de la recaptación de serotonina fluoxetina, fluvoxamina y sertralina para las algas". Ecotoxicología y seguridad ambiental . 67 (1): 128–139. Bibcode :2007EcoES..67..128J. doi :10.1016/j.ecoenv.2006.03.016. PMID 16753215.
- ^ McGowan K, Guerina V, Wicks J, Donowitz M (1985). "Hormonas secretoras de Entamoeba histolytica ". Simposio de la Fundación Ciba . Simposios de la Fundación Novartis. Vol. 112. págs. 139-154. doi :10.1002/9780470720936.ch8. ISBN 978-0-470-72093-6. Número de identificación personal 2861068.
- ^ Banu N, Zaidi KR, Mehdi G, Mansoor T (julio de 2005). "Alteraciones neurohumorales y su papel en la amebiasis". Revista india de bioquímica clínica . 20 (2): 142-145. doi :10.1007/BF02867414. PMC 3453840 . PMID 23105547.
- ^ Acharya DP, Sen MR, Sen PC (agosto de 1989). "Efecto de la 5-hidroxitriptamina exógena sobre la patogenicidad de Entamoeba histolytica en animales de experimentación". Indian Journal of Experimental Biology . 27 (8): 718–720. PMID 2561282.
- ^ Schröder P, Abele C, Gohr P, Stuhlfauth-Roisch U, Grosse W (1999). "Últimas novedades sobre la enzimología de la biosíntesis de serotonina en las semillas de nuez". Triptófano, serotonina y melatonina . Avances en medicina y biología experimental. Vol. 467. págs. 637–644. doi :10.1007/978-1-4615-4709-9_81. ISBN . 978-0-306-46204-7. Número de identificación personal 10721112.
- ^ Pénez N, Culioli G, Pérez T, Briand JF, Thomas OP, Blache Y (octubre de 2011). "Propiedades antiincrustantes de alcaloides indólicos y purínicos simples de la gorgonia mediterránea Paramuricea clavata". Revista de Productos Naturales . 74 (10): 2304–2308. doi :10.1021/np200537v. PMID 21939218.
- ^ Guillén-Casla V, Rosales-Conrado N, León-González ME, Pérez-Arribas LV, Polo-Díez LM (abril de 2012). "Determinación de serotonina y sus precursores en muestras de chocolate mediante cromatografía líquida capilar con detección por espectrometría de masas". Journal of Chromatography A . 1232 : 158–165. doi :10.1016/j.chroma.2011.11.037. PMID 22186492.
- ^ Pelagio-Flores R, Ortíz-Castro R, Méndez-Bravo A, Macías-Rodríguez L, López-Bucio J (marzo de 2011). "La serotonina, una señal derivada del triptófano conservada en plantas y animales, regula la arquitectura del sistema radicular probablemente actuando como un inhibidor natural de la auxina en Arabidopsis thaliana". Plant & Cell Physiology . 52 (3). Oxford University Press (OUP): 490–508. doi : 10.1093/pcp/pcr006 . PMID 21252298.
- ^ ab Powell JJ, Carere J, Fitzgerald TL, Stiller J, Covarelli L, Xu Q, et al. (marzo de 2017). "El patógeno de la podredumbre de la corona Fusarium pseudograminearum desencadena una serie de cambios transcripcionales y metabólicos en el trigo panificable (Triticum aestivum L.)". Anales de botánica . 119 (5). Oxford University Press (OUP): 853–867. doi : 10.1093/aob/mcw207 . PMC 5604588 . PMID 27941094. S2CID 3823345.
- ^ Du Fall LA, Solomon PS (octubre de 2013). "El efector necrotrófico SnToxA induce la síntesis de una nueva fitoalexina en el trigo". The New Phytologist . 200 (1). Wiley : 185–200. doi : 10.1111/nph.12356 . PMID 23782173.
- ^ Pasquet JC, Chaouch S, Macadré C, Balzergue S, Huguet S, Martin-Magniette ML, et al. (julio de 2014). "Expresión genética diferencial y análisis metabolómicos de Brachypodium distachyon infectado por cepas de Fusarium graminearum productoras y no productoras de deoxinivalenol". BMC Genomics . 15 (1). BioMed Central : 629. doi : 10.1186/1471-2164-15-629 . PMC 4124148 . PMID 25063396.
- ^ Zeiss DR, Piater LA, Dubery IA (febrero de 2021). "Amidas de hidroxicinamato: conjugados intrigantes de metabolitos protectores de plantas". Tendencias en la ciencia vegetal . 26 (2). Cell Press : 184–195. Bibcode :2021TPS....26..184Z. doi :10.1016/j.tplants.2020.09.011. PMID 33036915. S2CID 222256660.
- ^ Jonz MG, Ekaterini Mercier A, Joffre Potter JW (2001). "Efectos de la 5-HT (serotonina) en el comportamiento reproductivo en Heterodera Schachtii (Nematoda)". Revista Canadiense de Zoología . 79 (9): 1727. doi :10.1139/z01-135.
- ^ Sawin ER, Ranganathan R, Horvitz HR (junio de 2000). "La tasa locomotora de C. elegans está modulada por el entorno a través de una vía dopaminérgica y por la experiencia a través de una vía serotoninérgica". Neuron . 26 (3): 619–631. doi : 10.1016/S0896-6273(00)81199-X . PMID 10896158. S2CID 9247380.
- ^ Niacaris T, Avery L (enero de 2003). "La serotonina regula la repolarización del músculo faríngeo de C. elegans". The Journal of Experimental Biology . 206 (Pt 2): 223–231. doi :10.1242/jeb.00101. PMC 4441752 . PMID 12477893.
- ^ Rosso MN, Jones JT, Abad P (2009). "RNAi y genómica funcional en nematodos parásitos de plantas". Revisión anual de fitopatología . 47 (1). Revisiones anuales : 207–232. doi :10.1146/annurev.phyto.112408.132605. PMID 19400649. p. 218:
La octopamina y la serotonina regulan la actividad de las neuronas M3 que dirigen la contracción de la faringe durante la alimentación
de C. elegans
... La inmersión de
Meloidogyne
J2 en dsRNA en presencia de... resorcinol más serotonina resultó en la captación de soluciones y el silenciamiento de genes expresados en el intestino y las glándulas esofágicas.
- ^ Yeh SR, Fricke RA, Edwards DH (enero de 1996). "El efecto de la experiencia social en la modulación serotoninérgica del circuito de escape del cangrejo de río". Science . 271 (5247): 366–369. Bibcode :1996Sci...271..366Y. CiteSeerX 10.1.1.470.6528 . doi :10.1126/science.271.5247.366. PMID 8553075. S2CID 1575533.
- ^ Fry BG, Roelants K, Champagne DE, Scheib H, Tyndall JD, King GF, et al. (2009). "El multiverso toxicogenómico: reclutamiento convergente de proteínas en venenos animales". Revisión anual de genómica y genética humana . 10 (1). Revisiones anuales : 483–511. doi : 10.1146/annurev.genom.9.081307.164356 . PMID 19640225.
- ^ "Serotonina, receptores de serotonina y sus acciones en insectos". Neurotransmisor . 2 : 1–14. 2015. doi : 10.14800/nt.314 .
- ^ abc Schoofs L, De Loof A, Van Hiel MB (enero de 2017). "Neuropéptidos como reguladores del comportamiento en insectos". Revisión anual de entomología . 62 (1). Revisiones anuales : 35–52. doi : 10.1146/annurev-ento-031616-035500 . PMID 27813667.
- ^ ab Wang X, Kang L (7 de enero de 2014). "Mecanismos moleculares del cambio de fase en langostas". Revisión anual de entomología . 59 (1). Revisiones anuales : 225–244. doi : 10.1146/annurev-ento-011613-162019 . PMID 24160426.
p.
231,
El cambio en el número de varios neurotransmisores potenciales... como la serotonina... puede desempeñar un papel importante en la remodelación del SNC durante el cambio de fase (26, 56, 80).
p.
233,
En la langosta
S. gregaria
, la cantidad de serotonina en los ganglios torácicos se correlacionó positivamente con el grado de comportamiento gregario inducido por diferentes períodos de hacinamiento. Una serie de experimentos farmacológicos y conductuales demostraron que la serotonina desempeña un papel clave en la inducción de la gregarización conductual inicial (2, 80). Sin embargo, la serotonina no es responsable de mantener el comportamiento gregario porque su cantidad en langostas gregarias a largo plazo es menos de la mitad que en langostas solitarias a largo plazo (80). En
L. migratoria
, la inyección de serotonina también puede iniciar ligeramente el comportamiento gregario, pero la serotonina cuando acompaña al tratamiento de hacinamiento indujo un comportamiento más solitario que la inyección de serotonina sola (48). No se encontraron diferencias significativas en los niveles de serotonina en los tejidos cerebrales entre las dos fases de
L. migratoria
. Un informe reciente de Tanaka y Nishide (97) midió el comportamiento de atracción/evitación en
S. gregaria
después de inyecciones simples y múltiples de serotonina a diferentes concentraciones. La serotonina solo tuvo un efecto a corto plazo en el nivel de algunas actividades locomotoras y no estuvo involucrada en el control del comportamiento gregario (97). Además, no está claro cómo el neurotransmisor influye en este comportamiento único, porque un modelo de regresión logística binaria utilizado en estos estudios para el ensayo conductual se centró principalmente en un solo parámetro conductual que representa un estado de fase general. Obviamente, el cambio de fase conductual podría implicar mecanismos reguladores alternativos en diferentes especies de langostas. Por lo tanto, estos estudios demuestran que los mecanismos reguladores del SNC que rigen la iniciación y el mantenimiento del cambio de fase son específicos de cada especie e involucran las interacciones entre estos neurotransmisores.
Dados los papeles clave de la señalización aminérgica, ¿cuáles son las vías descendentes involucradas en el establecimiento de la memoria a largo plazo? Ott et al. (63) investigaron el papel de la proteína quinasa [] en el cambio de fase en
S. gregaria
: ... proteína quinasa A dependiente de AMPc (PKA). Mediante el uso de intervención farmacológica y de ARNip, estos autores han demostrado que la PKA... tiene un papel fundamental en la modulación de la propensión de las langostas a adquirir y expresar un comportamiento gregario. ... Desafortunadamente, aunque se planteó la hipótesis de una correlación entre la serotonina y la PKA, no se proporcionó evidencia directa. - ^ ab Zhang L, Lecoq M, Latchininsky A, Hunter D (enero de 2019). "Manejo de langostas y saltamontes". Revisión anual de entomología . 64 (1). Revisiones anuales : 15–34. doi : 10.1146/annurev-ento-011118-112500 . PMID 30256665. S2CID 52843907. p. 20:
...la gregarización es provocada por... estimulación táctil... Los estímulos táctiles desencadenan el aumento de aminas biógenas, particularmente serotonina, en el sistema nervioso de la langosta (1, 116); estas aminas desempeñan un papel crítico en la neurofisiología del cambio de fase del comportamiento de la langosta.
- ^ ab Anstey ML, Rogers SM, Ott SR, Burrows M, Simpson SJ (enero de 2009). "La serotonina media la gregarización conductual que subyace a la formación de enjambres en las langostas del desierto". Science . 323 (5914): 627–630. Bibcode :2009Sci...323..627A. doi :10.1126/science.1165939. PMID 19179529. S2CID 5448884.
- Morgan J (29 de enero de 2009). "Enjambres de langostas 'altos' de serotonina". BBC News .
- ^ Sitaraman D, LaFerriere H, Birman S, Zars T (junio de 2012). "La serotonina es fundamental para la memoria olfativa a corto plazo con recompensa en Drosophila". Journal of Neurogenetics . 26 (2): 238–244. doi :10.3109/01677063.2012.666298. PMID 22436011. S2CID 23639918.
- ^ Bicker G, Menzel R (enero de 1989). "Códigos químicos para el control del comportamiento en artrópodos". Nature . 337 (6202): 33–39. Bibcode :1989Natur.337...33B. doi :10.1038/337033a0. PMID 2562906. S2CID 223750.
- ^ Cai M, Li Z, Fan F, Huang Q, Shao X, Song G (marzo de 2010). "Diseño y síntesis de nuevos insecticidas basados en el ligando serotoninérgico 1-[(4-aminofenil)etil]-4-[3-(trifluorometil)fenil]piperazina (PAPP)". Journal of Agricultural and Food Chemistry . 58 (5): 2624–2629. doi :10.1021/jf902640u. PMID 20000410.
- ^ Manahan SE (2002). Química y bioquímica toxicológica (3.ª ed.). CRC Press. pág. 393. ISBN 978-1-4200-3212-3.
- ^ Postma TL (2009). "Venenos y venenos neurotóxicos para animales". En Dobbs MR (ed.). Neurotoxicología clínica . WB Saunders. págs. 463–489. doi :10.1016/B978-032305260-3.50049-6. ISBN 978-0-323-05260-3.
- ^ Gadenne C, Barrozo RB, Anton S (11 de marzo de 2016). "Plasticidad en el olfato de los insectos: ¿Oler o no oler?". Revista anual de entomología . 61 (1). Revistas anuales : 317–333. doi :10.1146/annurev-ento-010715-023523. hdl : 11336/19586 . PMID 26982441. S2CID 207568844.
- ^ Dierick HA, Greenspan RJ (mayo de 2007). "La serotonina y el neuropéptido F tienen efectos moduladores opuestos sobre la agresión de las moscas". Nature Genetics . 39 (5): 678–682. doi :10.1038/ng2029. PMID 17450142. S2CID 33768246.
- ^ Stoffolano JG , Haselton AT (7 de enero de 2013). "El buche de los dípteros adultos: un órgano único y pasado por alto". Revisión anual de entomología . 58 (1). Revisiones anuales : 205–225. doi :10.1146/annurev-ento-120811-153653. PMID 23317042.
- ^ abcdef Bacqué-Cazenave J, Bharatiya R, Barrière G, Delbecque JP, Bouguiyoud N, Di Giovanni G, et al. (febrero de 2020). "Serotonina en la cognición y el comportamiento animal". Revista Internacional de Ciencias Moleculares . 21 (5): 1649. doi : 10.3390/ijms21051649 . PMC 7084567 . PMID 32121267.
- ^ abcde Lucki I (agosto de 1998). "El espectro de conductas influidas por la serotonina". Psiquiatría biológica . 44 (3): 151–162. doi : 10.1016/s0006-3223(98)00139-5 . PMID 9693387. S2CID 3001666.
- ^ abc Backström T, Winberg S (25 de octubre de 2017). "La serotonina coordina las respuestas al estrés social: lo que podemos aprender de los peces". Frontiers in Neuroscience . 11 : 595. doi : 10.3389/fnins.2017.00595 . PMC 5669303 . PMID 29163002.
- ^ Berger M, Gray JA, Roth BL (1 de febrero de 2009). "La biología expandida de la serotonina". Revista Anual de Medicina . 60 (1): 355–366. doi :10.1146/annurev.med.60.042307.110802. PMC 5864293 . PMID 19630576.
- ^ Alberghina D, Piccione G, Pumilia G, Gioè M, Rizzo M, Raffo P, et al. (julio de 2019). "Fluctuación diaria de la serotonina y el cortisol en la orina en perros de refugio sanos e influencia de la exposición social intraespecífica". Fisiología y comportamiento . 206 : 1–6. doi :10.1016/j.physbeh.2019.03.016. PMID 30898540. S2CID 81965422.
- ^ Riggio G, Mariti C, Sergi V, Diverio S, Gazzano A (diciembre de 2020). "Concentraciones séricas de serotonina y triptófano en perros de refugio que muestran diferentes respuestas conductuales a un procedimiento potencialmente estresante". Ciencias Veterinarias . 8 (1): 1. doi : 10.3390/vetsci8010001 . PMC 7824451 . PMID 33374183.
- ^ Hydbring-Sandberg E, Larsson E, Madej A, Höglund OV (julio de 2021). "Efecto a corto plazo de la ovariohisterectomía sobre la serotonina, el cortisol, la testosterona y la progesterona en la orina de las perras". BMC Research Notes . 14 (1): 265. doi : 10.1186/s13104-021-05680-y . PMC 8272283 . PMID 34246304.
- ^ abc Winberg S, Thörnqvist PO (junio de 2016). "El papel de la serotonina cerebral en la modulación del comportamiento de los peces". Zoología actual . 62 (3): 317–323. doi :10.1093/cz/zow037. PMC 5804243 . PMID 29491919.
- ^ abcd Olivier B, Mos J, van der Heyden J, Hartog J (1 de febrero de 1989). "Modulación serotoninérgica de las interacciones sociales en ratones machos aislados". Psicofarmacología . 97 (2): 154–156. doi :10.1007/BF00442239. PMID 2498921. S2CID 37170174.
- ^ Huber R, Smith K, Delago A, Isaksson K, Kravitz EA (mayo de 1997). "Serotonina y motivación agresiva en crustáceos: alteración de la decisión de retirarse". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 94 (11): 5939–5942. Bibcode :1997PNAS...94.5939H. doi : 10.1073/pnas.94.11.5939 . PMC 20885 . PMID 9159179.
- ^ Sanchez CL, Biskup CS, Herpertz S, Gaber TJ, Kuhn CM, Hood SH, et al. (mayo de 2015). "El papel de la serotonina (5-HT) en el control del comportamiento: hallazgos de la investigación con animales e implicaciones clínicas". The International Journal of Neuropsychopharmacology . 18 (10): pyv050. doi :10.1093/ijnp/pyv050. PMC 4648158 . PMID 25991656.
- ^ abcd Petersen CL, Hurley LM (octubre de 2017). "Poniéndolo en contexto: vinculando el procesamiento auditivo con los circuitos de comportamiento social en el cerebro de los vertebrados". Biología comparativa e integradora . 57 (4): 865–877. doi :10.1093/icb/icx055. PMC 6251620 . PMID 28985384.
- ^ Loer CM, Kenyon CJ (diciembre de 1993). "Mutantes deficientes en serotonina y comportamiento de apareamiento masculino en el nematodo Caenorhabditis elegans". The Journal of Neuroscience . 13 (12): 5407–5417. doi :10.1523/JNEUROSCI.13-12-05407.1993. PMC 6576401 . PMID 8254383.
- ^ Lipton J, Kleemann G, Ghosh R, Lints R, Emmons SW (agosto de 2004). "Búsqueda de pareja en Caenorhabditis elegans: un modelo genético para el impulso sexual en un invertebrado simple". The Journal of Neuroscience . 24 (34): 7427–7434. doi :10.1523/JNEUROSCI.1746-04.2004. PMC 6729642 . PMID 15329389.
- ^ ab Murakami H, Murakami S (agosto de 2007). "Los receptores de serotonina modulan antagónicamente la longevidad de Caenorhabditis elegans". Aging Cell . 6 (4): 483–488. doi : 10.1111/j.1474-9726.2007.00303.x . PMID 17559503. S2CID 8345654.
- ^ Kaplan DD, Zimmermann G, Suyama K, Meyer T, Scott MP (julio de 2008). "Una GTPasa de la familia de nucleosteminas, NS3, actúa en neuronas serotoninérgicas para regular la señalización de insulina y controlar el tamaño corporal". Genes & Development . 22 (14): 1877–1893. doi :10.1101/gad.1670508. PMC 2492735 . PMID 18628395.
- ^ Ruaud AF, Thummel CS (julio de 2008). "La señalización de la serotonina y la insulina se unen para controlar el crecimiento en Drosophila". Genes & Development . 22 (14): 1851–1855. doi :10.1101/gad.1700708. PMC 2735276 . PMID 18628391.
- ^ Davidson S, Prokonov D, Taler M, Maayan R, Harell D, Gil-Ad I, et al. (febrero de 2009). "Efecto de la exposición a inhibidores selectivos de la recaptación de serotonina en el útero sobre el crecimiento fetal: posible función de los ejes IGF-I y HPA". Pediatric Research . 65 (2): 236–241. doi : 10.1203/PDR.0b013e318193594a . PMID 19262294.
- ^ Ben Arous J, Laffont S, Chatenay D (octubre de 2009). Brezina V (ed.). "Bases moleculares y sensoriales de un comportamiento de dos estados relacionado con los alimentos en C. elegans". PLOS ONE . 4 (10): e7584. Bibcode :2009PLoSO...4.7584B. doi : 10.1371/journal.pone.0007584 . PMC 2762077 . PMID 19851507.
- ^ Sze JY, Victor M, Loer C, Shi Y, Ruvkun G (febrero de 2000). "Defectos de señalización metabólica y alimentaria en un mutante de síntesis de serotonina de Caenorhabditis elegans". Naturaleza . 403 (6769): 560–564. Código Bib :2000Natur.403..560S. doi :10.1038/35000609. PMID 10676966. S2CID 4394553.
- ^ Murakami H, Bessinger K, Hellmann J, Murakami S (julio de 2008). "La manipulación de la señal de serotonina suprime la fase temprana del envejecimiento conductual en Caenorhabditis elegans". Neurobiología del envejecimiento . 29 (7): 1093–1100. doi :10.1016/j.neurobiolaging.2007.01.013. PMID 17336425. S2CID 37671716.
- ^ Côté F, Thévenot E, Fligny C, Fromes Y, Darmon M, Ripoche MA, et al. (noviembre de 2003). "La alteración del gen no neuronal tph1 demuestra la importancia de la serotonina periférica en la función cardíaca". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 100 (23): 13525–13530. Bibcode :2003PNAS..10013525C. doi : 10.1073/pnas.2233056100 . PMC 263847 . PMID 14597720.
- ^ Alarcón J, Cid E, Lillo L, Céspedesa C, Aguila S, Alderete JB (2008). "Biotransformación de derivados de indol por cultivos de micelio". Zeitschrift für Naturforschung C . 63 (1–2): 82–84. doi : 10.1515/znc-2008-1-215 . PMID 18386493. S2CID 29472174.
- ^ Sismaet HJ, Goluch ED (junio de 2018). "Sondas electroquímicas del comportamiento de la comunidad microbiana". Revisión anual de química analítica . 11 (1). Revisiones anuales : 441–461. Bibcode :2018ARAC...11..441S. doi : 10.1146/annurev-anchem-061417-125627 . PMID 29490192. S2CID 3632265. p. 449:
Tabla 1
Los picos de potencial respectivos para varias biomoléculas electroactivas que son producidas o consumidas por microbios reportados en la literatura
a
... Serotonina | Óxido de indio y estaño | +0,67 | 66
- ^ Henson JM, Butler MJ, Day AW (1999). "EL LADO OSCURO DEL MICELIO: Melaninas de hongos fitopatógenos". Revisión anual de fitopatología . 37 (1). Revisiones anuales : 447–471. doi :10.1146/annurev.phyto.37.1.447. PMID 11701831.
- ^ Anthony M (diciembre de 1984). "Antagonistas de la serotonina". Revista de Medicina de Australia y Nueva Zelanda . 14 (6): 888–895. doi :10.1111/j.1445-5994.1984.tb03802.x. PMID 6398056. S2CID 28327178.
- ^ Erspamer V (diciembre de 1954). "Farmacología de las indol-alquilaminas". Pharmacological Reviews . 6 (4): 425–487. PMID 13236482.
- ^ Negri L (2006). "[Vittorio Erspamer (1909-1999)]". Medicina Nei Secoli . 18 (1): 97-113. PMID 17526278.
- ^ Rapport MM, Green AA, Page IH (diciembre de 1948). "Serotonina, vasoconstrictor sérico; aislamiento y caracterización". The Journal of Biological Chemistry . 176 (3): 1243–1251. doi : 10.1016/S0021-9258(18)57137-4 . PMID 18100415.
- ^ Feldberg W, Toh CC (febrero de 1953). "Distribución de 5-hidroxitriptamina (serotonina, enteramina) en la pared del tracto digestivo". The Journal of Physiology . 119 (2–3): 352–362. doi :10.1113/jphysiol.1953.sp004850. PMC 1392800 . PMID 13035756.
- ^ SciFinder – Serotonin Substance Detail. Consultado el 4 de noviembre de 2012. [ Se necesita cita completa ]
- ^ Twarog BM, Page IH (octubre de 1953). "Contenido de serotonina en algunos tejidos y orina de mamíferos y un método para su determinación". The American Journal of Physiology . 175 (1): 157–161. doi : 10.1152/ajplegacy.1953.175.1.157 . PMID 13114371.
- ^ Página IH (julio de 1954). "Serotonina (5-hidroxitriptamina)". Physiological Reviews . 34 (3). American Physiological Society : 563–588. doi :10.1152/physrev.1954.34.3.563. PMID 13185755.
Lectura adicional
- Gutknecht L, Jacob C, Strobel A, Kriegebaum C, Müller J, Zeng Y, et al. (junio de 2007). "La variación del gen de la triptófano hidroxilasa-2 influye en los rasgos de personalidad y los trastornos relacionados con la desregulación emocional". Revista Internacional de Neuropsicofarmacología . 10 (3): 309–320. doi : 10.1017/S1461145706007437 . PMID 17176492.
Enlaces externos
- Espectro de espectrometría de masas de 5-hidroxitriptamina
- Serotonina unida a proteínas en el PDB
- PsychoTropicalResearch Amplias revisiones sobre fármacos serotoninérgicos y síndrome serotoninérgico.
- Molécula del mes: serotonina en la Universidad de Bristol
- 60-Second Psych: ¡No es justo! Mi nivel de serotonina es bajo, Scientific American
- Interpretación de la prueba de serotonina en ClinLab Navigator.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
