Lípido

Los lípidos son un amplio grupo de compuestos orgánicos que incluyen grasas , ceras , esteroles , vitaminas liposolubles (como las vitaminas A , D , E y K ), monoglicéridos , diglicéridos , fosfolípidos y otros. Las funciones de los lípidos incluyen almacenar energía, realizar señales y actuar como componentes estructurales de las membranas celulares . [3] [4] Los lípidos tienen aplicaciones en las industrias cosmética y alimentaria , y en la nanotecnología . [5]
Los lípidos pueden definirse ampliamente como moléculas pequeñas hidrofóbicas o anfifílicas ; la naturaleza anfifílica de algunos lípidos les permite formar estructuras como vesículas , liposomas multilamelares/unilamelares o membranas en un entorno acuoso. Los lípidos biológicos se originan total o parcialmente a partir de dos tipos distintos de subunidades bioquímicas o "bloques constructores": grupos cetoacilo e isopreno . [3] Usando este enfoque, los lípidos pueden dividirse en ocho categorías: acilos grasos , glicerolípidos , glicerofosfolípidos , esfingolípidos , sacarolípidos y policétidos (derivados de la condensación de subunidades cetoacilo); y lípidos de esterol y lípidos de prenol (derivados de la condensación de subunidades de isopreno). [3]
Aunque el término "lípido" se utiliza a veces como sinónimo de grasas, las grasas son un subgrupo de lípidos llamados triglicéridos . Los lípidos también abarcan moléculas como los ácidos grasos y sus derivados (incluidos los tri-, di-, monoglicéridos y fosfolípidos), así como otros metabolitos que contienen esteroles como el colesterol . [6] Aunque los humanos y otros mamíferos utilizan varias vías biosintéticas tanto para descomponer como para sintetizar lípidos, algunos lípidos esenciales no se pueden producir de esta manera y deben obtenerse de la dieta.
Historia
En 1815, Henri Braconnot clasificó los lípidos ( graisses ) en dos categorías, suifs (grasas sólidas o sebo) y huiles (aceites fluidos). [7] En 1823, Michel Eugène Chevreul desarrolló una clasificación más detallada, que incluía aceites, grasas, sebo, ceras, resinas, bálsamos y aceites volátiles (o aceites esenciales). [8] [9] [10]
El primer triglicérido sintético fue reportado por Théophile-Jules Pelouze en 1844, cuando produjo tributirina tratando ácido butírico con glicerina en presencia de ácido sulfúrico concentrado . [11] Varios años después, Marcellin Berthelot , uno de los estudiantes de Pelouze, sintetizó triestearina y tripalmitina por reacción de los ácidos grasos análogos con glicerina en presencia de cloruro de hidrógeno gaseoso a alta temperatura. [12]
En 1827, William Prout reconoció la grasa (materia alimenticia "oleosa"), junto con la proteína ("albúmina") y los carbohidratos ("sacarina"), como un nutriente importante para los humanos y los animales. [13] [14]
Durante un siglo, los químicos consideraron que las "grasas" eran simplemente lípidos compuestos de ácidos grasos y glicerol (glicéridos), pero más tarde se describieron nuevas formas. Theodore Gobley (1847) descubrió fosfolípidos en el cerebro de los mamíferos y en el huevo de gallina, a los que llamó " lecitinas ". Thudichum descubrió en el cerebro humano algunos fosfolípidos ( cefalina ), glicolípidos ( cerebrósido ) y esfingolípidos ( esfingomielina ). [9]
Los términos lipoide, lipina, lípido y lípido se han utilizado con significados variados de autor a autor. [15] En 1912, Rosenbloom y Gies propusieron la sustitución de "lipoide" por "lipina". [16] En 1920, Bloor introdujo una nueva clasificación para los "lipoides": lipoides simples (grasas y ceras), lipoides compuestos (fosfolipoides y glicolipoides) y los lipoides derivados (ácidos grasos, alcoholes , esteroles). [17] [18]
La palabra lípido , que proviene etimológicamente del griego λίπος, lipos 'grasa', fue introducida en 1923 por el farmacólogo francés Gabriel Bertrand . [19] Bertrand incluyó en el concepto no solo las grasas tradicionales (glicéridos), sino también los "lipoides", con una constitución compleja. [9] La palabra lípido fue aprobada por unanimidad por la comisión internacional de la Société de Chimie Biologique durante la sesión plenaria del 3 de julio de 1923. La palabra lípido fue posteriormente anglicanizada como lípido debido a su pronunciación ('lɪpɪd). En francés, el sufijo -ide , del griego antiguo -ίδης (que significa 'hijo de' o 'descendiente de'), siempre se pronuncia (ɪd).
En 1947, TP Hilditch definió los "lípidos simples" como grasas y ceras (ceras verdaderas, esteroles, alcoholes). [20] [ página necesaria ]
Categorías
El consorcio Lipid MAPS ha clasificado los lípidos en ocho categorías [3] de la siguiente manera:
Acilos grasos


Los acilos grasos, un término genérico para describir los ácidos grasos, sus conjugados y derivados, son un grupo diverso de moléculas sintetizadas por elongación de la cadena de un cebador de acetil-CoA con grupos malonil-CoA o metilmalonil-CoA en un proceso llamado síntesis de ácidos grasos . [21] [22] Están hechos de una cadena de hidrocarburo que termina con un grupo de ácido carboxílico ; esta disposición confiere a la molécula un extremo polar , hidrófilo , y un extremo no polar, hidrófobo que es insoluble en agua. La estructura de los ácidos grasos es una de las categorías más fundamentales de los lípidos biológicos y se utiliza comúnmente como un bloque de construcción de lípidos estructuralmente más complejos. La cadena de carbono, típicamente de entre cuatro y 24 carbonos de longitud, [23] puede estar saturada o insaturada , y puede estar unida a grupos funcionales que contienen oxígeno , halógenos , nitrógeno y azufre . Si un ácido graso contiene un doble enlace, existe la posibilidad de un isomería geométrica cis o trans , que afecta significativamente la configuración de la molécula . Los enlaces dobles cis hacen que la cadena de ácidos grasos se doble, un efecto que se agrava con más enlaces dobles en la cadena. Tres enlaces dobles en el ácido linolénico de 18 carbonos , las cadenas de acilo graso más abundantes en las membranas tilacoides de las plantas , hacen que estas membranas sean muy fluidas a pesar de las bajas temperaturas ambientales, [24] y también hacen que el ácido linolénico dé picos agudos dominantes en los espectros de RMN de 13-C de alta resolución de los cloroplastos. Esto a su vez juega un papel importante en la estructura y función de las membranas celulares. [25] : 193–5 La mayoría de los ácidos grasos naturales son de configuración cis , aunque la forma trans existe en algunas grasas y aceites naturales y parcialmente hidrogenados. [26]
Los ejemplos de ácidos grasos biológicamente importantes incluyen los eicosanoides , derivados principalmente del ácido araquidónico y el ácido eicosapentaenoico , que incluyen prostaglandinas , leucotrienos y tromboxanos . El ácido docosahexaenoico también es importante en los sistemas biológicos, particularmente con respecto a la vista. [27] [28] Otras clases importantes de lípidos en la categoría de ácidos grasos son los ésteres grasos y las amidas grasas. Los ésteres grasos incluyen intermediarios bioquímicos importantes como ésteres de cera , derivados de coenzima A de tioéster de ácido graso , derivados de ACP de tioéster de ácido graso y carnitinas de ácido graso. Las amidas grasas incluyen N-aciletanolaminas , como el neurotransmisor cannabinoide anandamida . [29]
Glicerolípidos

Los glicerolípidos están compuestos de gliceroles mono-, di- y tri-sustituidos , [30] siendo los más conocidos los triésteres de ácidos grasos del glicerol, llamados triglicéridos . La palabra "triacilglicerol" a veces se usa como sinónimo de "triglicérido". En estos compuestos, los tres grupos hidroxilo del glicerol están esterificados, típicamente por diferentes ácidos grasos. Debido a que funcionan como un depósito de energía, estos lípidos comprenden la mayor parte de la grasa almacenada en los tejidos animales. La hidrólisis de los enlaces éster de los triglicéridos y la liberación de glicerol y ácidos grasos del tejido adiposo son los pasos iniciales en la metabolización de la grasa. [31] : 630–1
Subclases adicionales de glicerolípidos están representadas por los glicosilgliceroles, que se caracterizan por la presencia de uno o más residuos de azúcar unidos al glicerol a través de un enlace glucosídico . Ejemplos de estructuras en esta categoría son los digalactosildiacilgliceroles que se encuentran en las membranas de las plantas [32] y los seminolípidos de los espermatozoides de los mamíferos [33] .
Glicerofosfolípidos

Los glicerofosfolípidos, generalmente denominados fosfolípidos (aunque las esfingomielinas también se clasifican como fosfolípidos), son ubicuos en la naturaleza y son componentes clave de la bicapa lipídica de las células, [34] además de estar involucrados en el metabolismo y la señalización celular . [35] El tejido neural (incluido el cerebro) contiene cantidades relativamente altas de glicerofosfolípidos, y las alteraciones en su composición se han implicado en varios trastornos neurológicos. [36] Los glicerofosfolípidos se pueden subdividir en clases distintas, según la naturaleza del grupo de cabeza polar en la posición sn -3 de la cadena principal de glicerol en eucariotas y eubacterias, o la posición sn -1 en el caso de las arqueobacterias . [37]
Ejemplos de glicerofosfolípidos encontrados en membranas biológicas son la fosfatidilcolina (también conocida como PC, GPCho o lecitina ), la fosfatidiletanolamina (PE o GPEtn) y la fosfatidilserina (PS o GPSer). Además de servir como un componente primario de las membranas celulares y sitios de unión para proteínas intra e intercelulares, algunos glicerofosfolípidos en células eucariotas, como los fosfatidilinositoles y los ácidos fosfatídicos son precursores o, en sí mismos, segundos mensajeros derivados de la membrana . [31] : 844 Por lo general, uno o ambos de estos grupos hidroxilo están acilados con ácidos grasos de cadena larga, pero también hay glicerofosfolípidos unidos a alquilo y unidos a 1Z-alquenilo ( plasmalógeno ), así como variantes de dialquiléter en arqueobacterias. [38]
Esfingolípidos

Los esfingolípidos son una familia compleja de compuestos [39] que comparten una característica estructural común, una estructura básica esfingoide que se sintetiza de novo a partir del aminoácido serina y un acil-CoA graso de cadena larga, y luego se convierte en ceramidas , fosfoesfingolípidos, glicoesfingolípidos y otros compuestos. La principal base esfingoide de los mamíferos se conoce comúnmente como esfingosina . Las ceramidas (bases N-acilo-esfingoideas) son una subclase importante de derivados de bases esfingoides con un ácido graso unido a amida . Los ácidos grasos son típicamente saturados o monoinsaturados con longitudes de cadena de 16 a 26 átomos de carbono. [25] : 421–2
Los principales fosfoesfingolípidos de los mamíferos son las esfingomielinas (fosfocolinas de ceramida), [40] mientras que los insectos contienen principalmente fosfoetanolaminas de ceramida [41] y los hongos tienen fosfoinositoles de fitoceramida y grupos de cabeza que contienen manosa . [42] Los glicoesfingolípidos son una familia diversa de moléculas compuestas de uno o más residuos de azúcar unidos a través de un enlace glucosídico a la base esfingoide. Ejemplos de estos son los glicoesfingolípidos simples y complejos como los cerebrósidos y los gangliósidos .
Esteroles
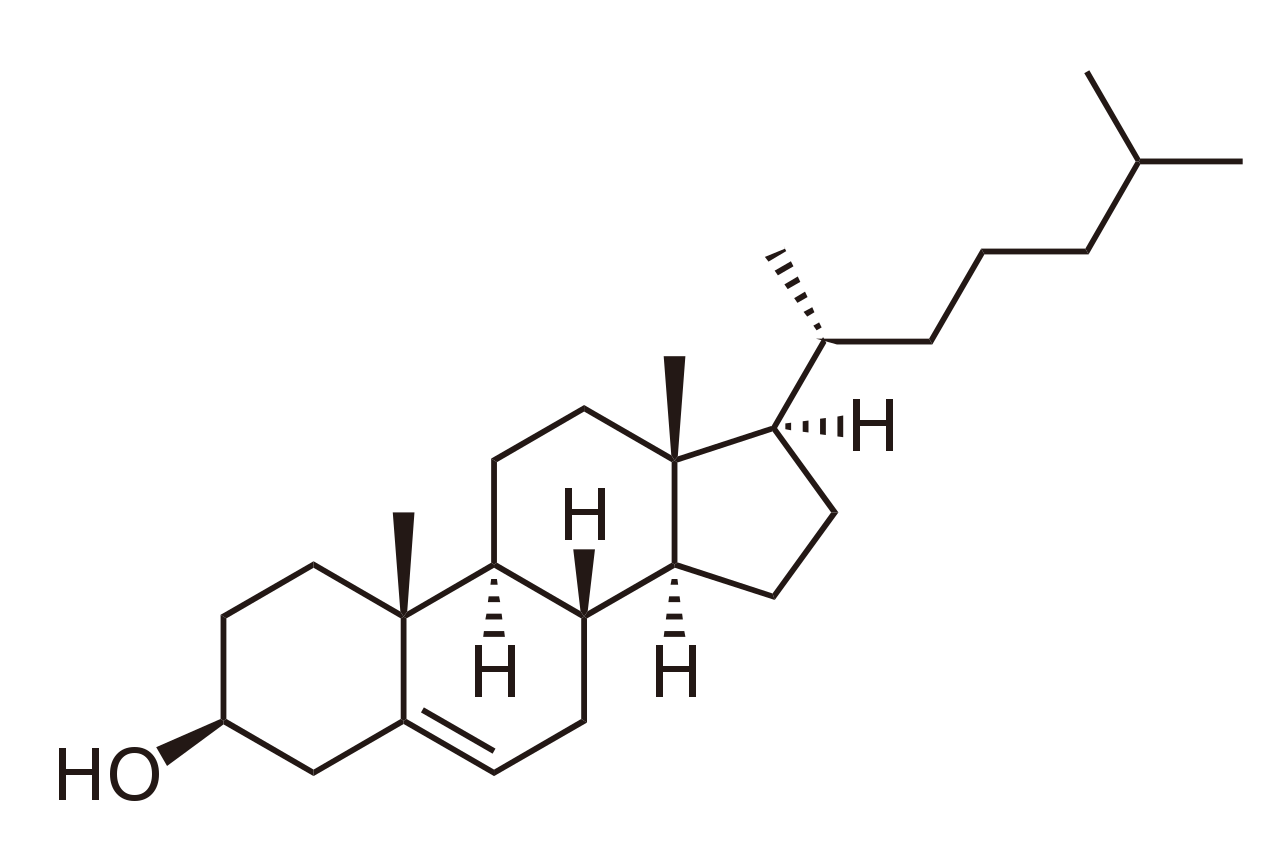
Los esteroles, como el colesterol y sus derivados, son un componente importante de los lípidos de membrana, [43] junto con los glicerofosfolípidos y las esfingomielinas. Otros ejemplos de esteroles son los ácidos biliares y sus conjugados, [44] que en los mamíferos son derivados oxidados del colesterol y se sintetizan en el hígado. Los equivalentes vegetales son los fitoesteroles , como el β-sitosterol , el estigmasterol y el brasicasterol ; este último compuesto también se utiliza como biomarcador del crecimiento de las algas . [45] El esterol predominante en las membranas celulares de los hongos es el ergosterol . [46]
Los esteroles son esteroides en los que uno de los átomos de hidrógeno está sustituido por un grupo hidroxilo , en la posición 3 de la cadena carbonada. Tienen en común con los esteroides la misma estructura central de cuatro anillos fusionados. Los esteroides tienen diferentes funciones biológicas como hormonas y moléculas de señalización . Los esteroides de dieciocho carbonos (C18) incluyen la familia de los estrógenos, mientras que los esteroides C19 comprenden los andrógenos, como la testosterona y la androsterona . La subclase C21 incluye los progestágenos , así como los glucocorticoides y mineralocorticoides . [2] : 749 Los secosteroides , que comprenden varias formas de vitamina D , se caracterizan por la escisión del anillo B de la estructura central. [47]
Prenoles

Los lípidos prenol se sintetizan a partir de los precursores de cinco unidades de carbono, el difosfato de isopentenilo y el difosfato de dimetilalilo , que se producen principalmente a través de la vía del ácido mevalónico (MVA). [48] Los isoprenoides simples (alcoholes lineales, difosfatos, etc.) se forman por la adición sucesiva de unidades C5 y se clasifican según el número de estas unidades de terpeno . Las estructuras que contienen más de 40 carbonos se conocen como politerpenos. Los carotenoides son isoprenoides simples importantes que funcionan como antioxidantes y como precursores de la vitamina A. [49] Otra clase biológicamente importante de moléculas está ejemplificada por las quinonas e hidroquinonas , que contienen una cola isoprenoide unida a un núcleo quinonoide de origen no isoprenoide. [50] La vitamina E y la vitamina K , así como las ubiquinonas , son ejemplos de esta clase. Los procariotas sintetizan poliprenoles (llamados bactoprenoles ) en los que la unidad isoprenoide terminal unida al oxígeno permanece insaturada, mientras que en los poliprenoles animales ( dolicoles ) el isoprenoide terminal se reduce. [51]
Sacarolípidos

Los sacarolípidos describen compuestos en los que los ácidos grasos están unidos a una cadena principal de azúcar, formando estructuras que son compatibles con las bicapas de membrana. En los sacarolípidos, un monosacárido sustituye a la cadena principal de glicerol presente en los glicerolípidos y glicerofosfolípidos. Los sacarolípidos más conocidos son los precursores de glucosamina acilada del componente Lípido A de los lipopolisacáridos en bacterias Gram-negativas . Las moléculas típicas de lípido A son disacáridos de glucosamina, que se derivatizan con hasta siete cadenas de acilo graso. El lipopolisacárido mínimo requerido para el crecimiento en E. coli es Kdo 2 -Lípido A, un disacárido hexaacilado de glucosamina que está glicosilado con dos residuos de ácido 3-desoxi-D-mano-octulosónico (Kdo). [52]
Policétidos
Los policétidos se sintetizan por polimerización de subunidades de acetilo y propionilo por enzimas clásicas, así como por enzimas iterativas y multimodulares que comparten características mecanísticas con las sintasas de ácidos grasos . Comprenden muchos metabolitos secundarios y productos naturales de fuentes animales, vegetales, bacterianas, fúngicas y marinas, y tienen una gran diversidad estructural. [53] [54] Muchos policétidos son moléculas cíclicas cuyas cadenas principales a menudo se modifican aún más por glicosilación , metilación , hidroxilación , oxidación u otros procesos. Muchos agentes antimicrobianos , antiparasitarios y anticancerígenos de uso común son policétidos o derivados de policétidos, como eritromicinas , tetraciclinas , avermectinas y epotilonas antitumorales . [55]
Funciones biológicas
Componente de las membranas biológicas
Las células eucariotas presentan orgánulos compartimentados unidos a membranas que llevan a cabo diferentes funciones biológicas. Los glicerofosfolípidos son el principal componente estructural de las membranas biológicas , como la membrana plasmática celular y las membranas intracelulares de los orgánulos; en las células animales, la membrana plasmática separa físicamente los componentes intracelulares del entorno extracelular . [ cita requerida ] Los glicerofosfolípidos son moléculas anfipáticas (que contienen regiones hidrófobas e hidrófilas) que contienen un núcleo de glicerol unido a dos "colas" derivadas de ácidos grasos por enlaces éster y a un grupo "cabeza" por un enlace éster fosfato . [ cita requerida ] Si bien los glicerofosfolípidos son el componente principal de las membranas biológicas, otros componentes lipídicos no glicéridos como la esfingomielina y los esteroles (principalmente el colesterol en las membranas celulares animales) también se encuentran en las membranas biológicas. [56] [2] : 329–331 En plantas y algas, los galactosildiacilgliceroles, [57] y el sulfoquinovosildiacilglicerol, [32] que carecen de un grupo fosfato, son componentes importantes de las membranas de los cloroplastos y orgánulos relacionados y se encuentran entre los lípidos más abundantes en los tejidos fotosintéticos, incluidos los de plantas superiores, algas y ciertas bacterias. [58]
Las membranas tilacoides de las plantas tienen el componente lipídico más grande de un monogalactosil diglicérido no formador de bicapa (MGDG) y pocos fosfolípidos; a pesar de esta composición lipídica única, se ha demostrado que las membranas tilacoides del cloroplasto contienen una matriz lipídica bicapa dinámica, como lo revelan los estudios de resonancia magnética y microscopio electrónico. [59]

Una membrana biológica es una forma de bicapa lipídica de fase laminar . La formación de bicapas lipídicas es un proceso energéticamente preferido cuando los glicerofosfolípidos descritos anteriormente están en un entorno acuoso. [2] : 333–4 Esto se conoce como efecto hidrofóbico . En un sistema acuoso, las cabezas polares de los lípidos se alinean hacia el entorno polar acuoso, mientras que las colas hidrofóbicas minimizan su contacto con el agua y tienden a agruparse, formando una vesícula ; dependiendo de la concentración del lípido, esta interacción biofísica puede resultar en la formación de micelas , liposomas o bicapas lipídicas . También se observan otras agregaciones que forman parte del polimorfismo del comportamiento anfifílico (lípido). El comportamiento de fase es un área de estudio dentro de la biofísica . [60] [61] Las micelas y las bicapas se forman en el medio polar mediante un proceso conocido como efecto hidrofóbico. [62] Al disolver una sustancia lipofílica o anfifílica en un entorno polar, las moléculas polares (es decir, el agua en una solución acuosa) se vuelven más ordenadas alrededor de la sustancia lipofílica disuelta, ya que las moléculas polares no pueden formar enlaces de hidrógeno con las áreas lipofílicas del anfifílico. Por lo tanto, en un entorno acuoso, las moléculas de agua forman una jaula de " clatrato " ordenada alrededor de la molécula lipofílica disuelta. [63]
La formación de lípidos en las membranas de las protocélulas representa un paso clave en los modelos de abiogénesis , el origen de la vida. [64]
Almacenamiento de energía
Los triglicéridos, almacenados en el tejido adiposo, son una forma importante de almacenamiento de energía tanto en animales como en plantas. Son una fuente importante de energía en la respiración aeróbica. La oxidación completa de los ácidos grasos libera alrededor de 38 kJ/g (9 kcal/g ), en comparación con solo 17 kJ/g (4 kcal/g) para la descomposición oxidativa de carbohidratos y proteínas . El adipocito , o célula grasa, está diseñado para la síntesis y descomposición continua de triglicéridos en animales, con una descomposición controlada principalmente por la activación de la enzima sensible a las hormonas, la lipasa . [65] Las aves migratorias que deben volar largas distancias sin comer usan triglicéridos para alimentar sus vuelos. [2] : 619
Señalización
Han surgido evidencias que muestran que la señalización lipídica es una parte vital de la señalización celular . [66] [67] [68] [69] La señalización lipídica puede ocurrir a través de la activación de receptores nucleares o acoplados a proteína G , y se han identificado miembros de varias categorías lipídicas diferentes como moléculas de señalización y mensajeros celulares . [70] Estos incluyen esfingosina-1-fosfato , un esfingolípido derivado de la ceramida que es una potente molécula mensajera involucrada en la regulación de la movilización de calcio, [71] el crecimiento celular y la apoptosis; [72] diacilglicerol y los fosfatos de fosfatidilinositol (PIP), involucrados en la activación mediada por calcio de la proteína quinasa C ; [73] las prostaglandinas , que son un tipo de eicosanoide derivado de ácidos grasos involucrado en la inflamación y la inmunidad ; [74] las hormonas esteroides como el estrógeno , la testosterona y el cortisol , que modulan una serie de funciones como la reproducción, el metabolismo y la presión arterial; y los oxiesteroles como el 25-hidroxi-colesterol que son agonistas del receptor X del hígado . [75] Se sabe que los lípidos de fosfatidilserina están involucrados en la señalización para la fagocitosis de células apoptóticas o fragmentos de células. Logran esto al estar expuestos a la cara extracelular de la membrana celular después de la inactivación de las flipasas que las colocan exclusivamente en el lado citosólico y la activación de las escramblasas, que alteran la orientación de los fosfolípidos. Después de que esto ocurre, otras células reconocen las fosfatidilserinas y fagocitan las células o fragmentos de células exponiéndolas. [76]
Otras funciones
Las vitaminas "liposolubles" ( A , D , E y K ), que son lípidos basados en isopreno , son nutrientes esenciales almacenados en el hígado y los tejidos grasos, con una amplia gama de funciones. Las acil-carnitinas están involucradas en el transporte y metabolismo de los ácidos grasos dentro y fuera de las mitocondrias, donde sufren oxidación beta . [77] Los poliprenoles y sus derivados fosforilados también juegan papeles importantes en el transporte, en este caso el transporte de oligosacáridos a través de las membranas. Los azúcares de fosfato de poliprenol y los azúcares de difosfato de poliprenol funcionan en reacciones de glicosilación extracitoplasmática, en la biosíntesis de polisacáridos extracelulares (por ejemplo, polimerización de peptidoglicano en bacterias) y en la N- glicosilación de proteínas eucariotas . [78] [79] Las cardiolipinas son una subclase de glicerofosfolípidos que contienen cuatro cadenas de acilo y tres grupos de glicerol que son particularmente abundantes en la membrana mitocondrial interna. [80] [81] Se cree que activan las enzimas involucradas en la fosforilación oxidativa . [82] Los lípidos también forman la base de las hormonas esteroides. [83]
Metabolismo
Los principales lípidos de la dieta de los seres humanos y otros animales son los triglicéridos animales y vegetales, los esteroles y los fosfolípidos de membrana. El proceso del metabolismo lipídico sintetiza y degrada los depósitos lipídicos y produce los lípidos estructurales y funcionales característicos de los tejidos individuales.
Biosíntesis
En los animales, cuando hay un exceso de carbohidratos en la dieta, el exceso de carbohidratos se convierte en triglicéridos. Esto implica la síntesis de ácidos grasos a partir de acetil-CoA y la esterificación de ácidos grasos en la producción de triglicéridos, un proceso llamado lipogénesis . [2] : 634 Los ácidos grasos son producidos por sintasas de ácidos grasos que polimerizan y luego reducen las unidades de acetil-CoA. Las cadenas de acilo en los ácidos grasos se extienden mediante un ciclo de reacciones que agregan el grupo acetilo, lo reducen a un alcohol, lo deshidratan a un grupo alqueno y luego lo reducen nuevamente a un grupo alcano . Las enzimas de la biosíntesis de ácidos grasos se dividen en dos grupos, en animales y hongos todas estas reacciones de sintasa de ácidos grasos son llevadas a cabo por una sola proteína multifuncional, [84] mientras que en los plástidos vegetales y las bacterias enzimas separadas realizan cada paso en la vía. [85] [86] Los ácidos grasos pueden convertirse posteriormente en triglicéridos que se empaquetan en lipoproteínas y se secretan desde el hígado.
La síntesis de ácidos grasos insaturados implica una reacción de desaturación , mediante la cual se introduce un doble enlace en la cadena de acilo graso. Por ejemplo, en los seres humanos, la desaturación del ácido esteárico por la estearoil-CoA desaturasa-1 produce ácido oleico . El ácido graso doblemente insaturado, el ácido linoleico , así como el ácido α-linolénico triplemente insaturado , no pueden sintetizarse en los tejidos de los mamíferos y, por lo tanto, son ácidos grasos esenciales y deben obtenerse de la dieta. [2] : 643
La síntesis de triglicéridos se lleva a cabo en el retículo endoplásmico mediante vías metabólicas en las que los grupos acilo de los acil-CoA grasos se transfieren a los grupos hidroxilo del glicerol-3-fosfato y el diacilglicerol. [2] : 733–9
Los terpenos e isoprenoides , incluidos los carotenoides , se forman mediante el ensamblaje y la modificación de unidades de isopreno donadas a partir de los precursores reactivos pirofosfato de isopentenilo y pirofosfato de dimetilalilo . [48] Estos precursores se pueden producir de diferentes maneras. En animales y arqueas , la vía del mevalonato produce estos compuestos a partir de acetil-CoA, [87] mientras que en plantas y bacterias la vía no mevalonato utiliza piruvato y gliceraldehído 3-fosfato como sustratos. [48] [88] Una reacción importante que utiliza estos donantes de isopreno activados es la biosíntesis de esteroides . Aquí, las unidades de isopreno se unen para formar escualeno y luego se pliegan y forman un conjunto de anillos para formar lanosterol . [89] El lanosterol luego se puede convertir en otros esteroides como el colesterol y el ergosterol. [89] [90]
Degradación
La beta oxidación es el proceso metabólico por el cual los ácidos grasos se descomponen en las mitocondrias o en los peroxisomas para generar acetil-CoA . En su mayor parte, los ácidos grasos se oxidan mediante un mecanismo que es similar, pero no idéntico, a una inversión del proceso de síntesis de ácidos grasos. Es decir, los fragmentos de dos carbonos se eliminan secuencialmente del extremo carboxilo del ácido después de los pasos de deshidrogenación , hidratación y oxidación para formar un beta-cetoácido , que se divide por tiolisis . El acetil-CoA se convierte finalmente en trifosfato de adenosina (ATP), CO2 y H2O utilizando el ciclo del ácido cítrico y la cadena de transporte de electrones . Por lo tanto, el ciclo del ácido cítrico puede comenzar en acetil-CoA cuando la grasa se está descomponiendo para obtener energía si hay poca o ninguna glucosa disponible. El rendimiento energético de la oxidación completa del palmitato de ácido graso es de 106 ATP. [2] : 625–6 Los ácidos grasos insaturados y de cadena impar requieren pasos enzimáticos adicionales para su degradación.
Nutrición y salud
La mayor parte de la grasa que se encuentra en los alimentos está en forma de triglicéridos, colesterol y fosfolípidos. Algunas grasas de la dieta son necesarias para facilitar la absorción de vitaminas liposolubles ( A , D , E y K ) y carotenoides . [91] : 903 Los humanos y otros mamíferos tienen un requerimiento dietético de ciertos ácidos grasos esenciales, como el ácido linoleico (un ácido graso omega-6 ) y el ácido alfa-linolénico (un ácido graso omega-3) porque no pueden sintetizarse a partir de precursores simples en la dieta. [2] : 643 Ambos ácidos grasos son ácidos grasos poliinsaturados de 18 carbonos que difieren en el número y la posición de los dobles enlaces. La mayoría de los aceites vegetales son ricos en ácido linoleico ( aceites de cártamo , girasol y maíz ). El ácido alfa-linolénico se encuentra en las hojas verdes de las plantas y en algunas semillas, nueces y legumbres (en particular lino , colza , nuez y soja ). [92] Los aceites de pescado son particularmente ricos en ácidos grasos omega-3 de cadena larga, ácido eicosapentaenoico y ácido docosahexaenoico . [91] : 388 Muchos estudios han demostrado beneficios positivos para la salud asociados con el consumo de ácidos grasos omega-3 en el desarrollo infantil, el cáncer, las enfermedades cardiovasculares y varias enfermedades mentales (como la depresión, el trastorno por déficit de atención con hiperactividad y la demencia). [93] [94]
Por el contrario, ahora está bien establecido que el consumo de grasas trans , como las presentes en los aceites vegetales parcialmente hidrogenados , es un factor de riesgo de enfermedad cardiovascular . Las grasas que son buenas para uno pueden convertirse en grasas trans por métodos de cocción inadecuados que dan como resultado una cocción excesiva de los lípidos. [95] [96] [97]
Algunos estudios han sugerido que la ingesta total de grasas en la dieta está relacionada con un mayor riesgo de obesidad. [98] [99] y diabetes; [100] Otros, incluido el Women's Health Initiative Dietary Modification Trial, un estudio de ocho años de 49.000 mujeres, el Nurses' Health Study y el Health Professionals Follow-up Study, no revelaron tales vínculos. [101] [102] Ninguno de estos estudios sugirió ninguna conexión entre el porcentaje de calorías provenientes de grasas y el riesgo de cáncer, enfermedades cardíacas o aumento de peso. The Nutrition Source, [103] un sitio web mantenido por el departamento de nutrición de la TH Chan School of Public Health de la Universidad de Harvard , resume la evidencia actual sobre el efecto de las grasas en la dieta: "Investigaciones detalladas, gran parte de ellas realizadas en Harvard, muestran que la cantidad total de grasas en la dieta no está realmente relacionada con el peso o las enfermedades". [104]
Véase también
- Nanopartículas lipídicas sólidas : un nuevo sistema de administración de fármacos
- Lípido simple
- Prueba de emulsión
- Microdominio lipídico
- Lípidos de membrana : moléculas lipídicas en la membrana celular
- Lipidómica : estudio a gran escala del metabolismo lipídico de un organismo utilizando técnicas de análisis químico de alta tecnología.Pages displaying wikidata descriptions as a fallback
- Lipidoma – Conjunto de lípidos en las células
- Interacción proteína-lípido
- Lípido fenólico : Clase de compuestos orgánicos, una clase de productos naturales compuestos de largas cadenas alifáticas y anillos fenólicos que se encuentran en plantas, hongos y bacterias.
Referencias
- ^ Maitland J Jr (1998). Química orgánica . WW Norton & Co Inc (Np). pág. 139. ISBN 978-0-393-97378-5.
- ^ abcdefghij Stryer L, Berg JM, Tymoczko JL (2007). Bioquímica (6ª ed.). San Francisco: WH Freeman. ISBN 978-0-7167-8724-2.
- ^ abcd Fahy E, Subramaniam S, Murphy RC, Nishijima M, Raetz CR, Shimizu T, Spener F, van Meer G, Wakelam MJ, Dennis EA (abril de 2009). "Actualización del sistema de clasificación integral LIPID MAPS para lípidos". Journal of Lipid Research . 50 (S1): S9–14. doi : 10.1194/jlr.R800095-JLR200 . PMC 2674711 . PMID 19098281.
- ^ Subramaniam S, Fahy E, Gupta S, Sud M, Byrnes RW, Cotter D, Dinasarapu AR, Maurya MR (octubre de 2011). "Bioinformática y biología de sistemas del lipidoma". Reseñas químicas . 111 (10): 6452–6490. doi :10.1021/cr200295k. PMC 3383319 . PMID 21939287.
- ^ Mashaghi S, Jadidi T, Koenderink G , Mashaghi A (febrero de 2013). "Nanotecnología lipídica". Revista internacional de ciencias moleculares . 14 (2): 4242–4282. doi : 10.3390/ijms14024242 . PMC 3588097. PMID 23429269 .

- ^ Michelle A, Hopkins J, McLaughlin CW, Johnson S, Warner MQ, LaHart D, Wright JD (1993). Biología humana y salud. Englewood Cliffs, Nueva Jersey: Prentice Hall. ISBN 978-0-13-981176-0.
- ^ Braconnot H (31 de marzo de 1815). "Sobre la naturaleza del cuerpo graso". Anales de chimenea . 2 (XCIII): 225–277.
- ^ Chevreul ME (1823). Recherches sur les corps gras d'origine animale. París: Levrault.
- ^ abc Leray C (2012). Introducción a la lipidómica. Boca Raton: CRC Press. ISBN 978-1466551466.
- ^ Leray C (2015). "Introducción, historia y evolución". Lípidos. Nutrición y salud. Boca Raton: CRC Press. ISBN 978-1482242317.
- ^ Pelouze TJ, Gélis A (1844). "Mémoire sur l'acide butyrique". Annales de Chimie et de Physique . 10 : 434.
- ^ Comptes rendus hebdomadaires des séances de l'Académie des Sciences, París, 1853, 36, 27; Annales de Chimie et de Physique 1854, 41, 216
- ^ Leray C. «Historia cronológica del centro lipídico». Cyberlipid Center . Archivado desde el original el 13 de octubre de 2017. Consultado el 1 de diciembre de 2017 .
- ^ Prout W (1827). "Sobre la composición última de sustancias alimenticias simples, con algunas observaciones preliminares sobre el análisis de cuerpos organizados en general". Phil. Trans. : 355–388.
- ^ Culling CF (1974). "Lípidos. (Grasas, Lipoides, Lipinas)". Manual de técnicas histopatológicas (3.ª ed.). Londres: Butterworths. págs. 351–376. ISBN 978-1483164793.
- ^ Rosenbloom J, Gies WJ (1911). "Sugerencia para profesores de bioquímica. I. Una propuesta de clasificación química de las lipinas, con una nota sobre la íntima relación entre los colesteroles y las sales biliares". Biochem. Bull . 1 : 51–56.
- ^ Bloor WR (1920). "Esquema de una clasificación de los lípidos". Proc. Soc. Exp. Biol. Med . 17 (6): 138–140. doi :10.3181/00379727-17-75. S2CID 75844378.
- ^ Christie WW, Han X (2010). Análisis de lípidos: aislamiento, separación, identificación y análisis lipidómico. Bridgwater, Inglaterra: The Oily Press. ISBN 978-0857097866.
- ^ Bertrand G (1923). "Proyecto de reforma de la nomenclatura de chimenea biológica". Boletín de la Sociedad de Chimie Biologique . 5 : 96-109.
- ^ Hilditch, Thomas Percy (1956). La constitución química de las grasas naturales. Wiley.
- ^ Vance JE, Vance DE (2002). Bioquímica de lípidos, lipoproteínas y membranas . Ámsterdam: Elsevier. ISBN 978-0-444-51139-3.
- ^ Brown HA, ed. (2007). Lipodomía y lípidos bioactivos: análisis de lípidos basado en espectrometría de masas . Métodos en enzimología. Vol. 423. Boston: Academic Press. ISBN 978-0-12-373895-0.
- ^ Hunt SM, Groff JL, Gropper SA (1995). Nutrición avanzada y metabolismo humano. Belmont, California: West Pub. Co. pág. 98. ISBN 978-0-314-04467-9.
- ^ Yashroy RC (1987). "Estudios de RMN de 13C de cadenas de acilo graso lipídico de membranas de cloroplastos". Indian Journal of Biochemistry and Biophysics . 24 (6): 177–178. doi :10.1016/0165-022X(91)90019-S. PMID 3428918.
- ^ ab Devlin TM (1997). Libro de texto de bioquímica: con correlaciones clínicas (4.ª ed.). Chichester: John Wiley & Sons. ISBN 978-0-471-17053-2.
- ^ Hunter JE (noviembre de 2006). "Ácidos grasos trans en la dieta: revisión de estudios recientes en humanos y respuestas de la industria alimentaria". Lípidos . 41 (11): 967–992. doi :10.1007/s11745-006-5049-y. PMID 17263298. S2CID 1625062.
- ^ Furse S (2 de diciembre de 2011). "Un lípido largo, un nombre largo: ácido docosahexaenoico". The Lipid Chronicles .
- ^ "DHA para un funcionamiento cerebral y visual óptimo". Instituto DHA/EPA Omega-3.
- ^ Fezza F, De Simone C, Amadio D, Maccarrone M (2008). "Amiduro hidrolasa de ácidos grasos: un guardián del sistema endocannabinoide". Lípidos en la salud y la enfermedad . Bioquímica subcelular. Vol. 49. págs. 101–132. doi :10.1007/978-1-4020-8831-5_4. ISBN 978-1-4020-8830-8. Número de identificación personal 18751909.
- ^ Coleman RA, Lee DP (marzo de 2004). "Enzimas de la síntesis de triacilglicerol y su regulación". Progress in Lipid Research . 43 (2): 134–176. doi :10.1016/S0163-7827(03)00051-1. PMID 14654091.
- ^ de van Holde KE, Mathews CK (1996). Bioquímica (2.ª ed.). Menlo Park, California: Benjamin/Cummings Pub. Co. ISBN 978-0-8053-3931-4.
- ^ ab Hölzl G, Dörmann P (septiembre de 2007). "Estructura y función de los glicoglicerolípidos en plantas y bacterias". Progreso en la investigación de lípidos . 46 (5): 225–243. doi :10.1016/j.plipres.2007.05.001. PMID 17599463.
- ^ Honke K, Zhang Y, Cheng X, Kotani N, Taniguchi N (2004). "Funciones biológicas de los sulfoglicolípidos y fisiopatología de su deficiencia". Glycoconjugate Journal . 21 (1–2): 59–62. doi :10.1023/B:GLYC.0000043749.06556.3d. PMID 15467400. S2CID 2678053.
- ^ "La estructura de una membrana". The Lipid Chronicles . 5 de noviembre de 2011. Consultado el 31 de diciembre de 2011 .
- ^ Berridge MJ, Irvine RF (septiembre de 1989). "Fosfatos de inositol y señalización celular". Nature . 341 (6239): 197–205. Bibcode :1989Natur.341..197B. doi :10.1038/341197a0. PMID 2550825. S2CID 26822092.
- ^ Farooqui AA, Horrocks LA, Farooqui T (junio de 2000). "Glicerofosfolípidos en el cerebro: su metabolismo, incorporación a las membranas, funciones y participación en trastornos neurológicos". Química y física de los lípidos . 106 (1): 1–29. doi :10.1016/S0009-3084(00)00128-6. PMID 10878232.
- ^ Ivanova PT, Milne SB, Byrne MO, Xiang Y, Brown HA (2007). "Identificación y cuantificación de glicerofosfolípidos mediante espectrometría de masas de ionización por electrospray". Lipidomics and Bioactive Lipids: Mass-Spectrometry–Based Lipid Analysis . Métodos en enzimología. Vol. 432. págs. 21–57. doi :10.1016/S0076-6879(07)32002-8. ISBN 978-0-12-373895-0. Número de identificación personal 17954212.
- ^ Paltauf F (diciembre de 1994). "Lípidos de éter en biomembranas". Química y física de lípidos . 74 (2): 101–139. doi :10.1016/0009-3084(94)90054-X. PMID 7859340.
- ^ Merrill AH, Sandoff K (2002). "Capítulo 14: Esfingolípidos: metabolismo y señalización celular" (PDF) . En Vance JE, Vance EE (eds.). Bioquímica de lípidos, lipoproteínas y membranas (4.ª ed.). Ámsterdam: Elsevier. págs. 373–407. ISBN 978-0-444-51138-6.
- ^ Hori T, Sugita M (1993). "Esfingolípidos en animales inferiores". Progreso en la investigación de lípidos . 32 (1): 25–45. doi :10.1016/0163-7827(93)90003-F. PMID 8415797.
- ^ Wiegandt H (enero de 1992). "Glicolípidos de insectos". Biochimica et Biophysica Acta (BBA) - Lípidos y metabolismo de los lípidos . 1123 (2): 117–126. doi :10.1016/0005-2760(92)90101-Z. PMID 1739742.
- ^ Guan X, Wenk MR (mayo de 2008). "Bioquímica de los lípidos de inositol". Frontiers in Bioscience . 13 (13): 3239–3251. doi : 10.2741/2923 . PMID 18508430.
- ^ Bach D, Wachtel E (marzo de 2003). "Membranas modelo de fosfolípidos/colesterol: formación de cristales de colesterol". Biochimica et Biophysica Acta (BBA) - Biomembranas . 1610 (2): 187–197. doi : 10.1016/S0005-2736(03)00017-8 . PMID 12648773.
- ^ Russell DW (2003). "Las enzimas, la regulación y la genética de la síntesis de ácidos biliares". Revista anual de bioquímica . 72 : 137–174. doi :10.1146/annurev.biochem.72.121801.161712. PMID 12543708.
- ^ Villinski JC, Hayes JM, Brassell SC, Riggert VL, Dunbar R (2008). "Esteroles sedimentarios como indicadores biogeoquímicos en el Océano Austral". Geoquímica orgánica . 39 (5): 567–588. Código Bibliográfico :2008OrGeo..39..567V. doi :10.1016/j.orggeochem.2008.01.009.
- ^ Deacon J (2005). Biología de los hongos . Cambridge, Massachusetts: Blackwell Publishers. pág. 342. ISBN. 978-1-4051-3066-0.
- ^ Bouillon R, Verstuyf A, Mathieu C, Van Cromphaut S, Masuyama R, Dehaes P, Carmeliet G (diciembre de 2006). "Resistencia a la vitamina D". Mejores prácticas e investigación. Endocrinología clínica y metabolismo . 20 (4): 627–645. doi :10.1016/j.beem.2006.09.008. PMID 17161336.
- ^ abc Kuzuyama T, Seto H (abril de 2003). "Diversidad de la biosíntesis de las unidades de isopreno". Natural Product Reports . 20 (2): 171–183. doi :10.1039/b109860h. PMID 12735695.
- ^ Rao AV, Rao LG (marzo de 2007). "Carotenoides y salud humana". Investigación farmacológica . 55 (3): 207–216. doi :10.1016/j.phrs.2007.01.012. PMID 17349800.
- ^ Brunmark A, Cadenas E (1989). "Química redox y de adición de compuestos quinoides y sus implicaciones biológicas". Free Radical Biology & Medicine . 7 (4): 435–477. doi :10.1016/0891-5849(89)90126-3. PMID 2691341.
- ^ Swiezewska E, Danikiewicz W (julio de 2005). "Poliisoprenoides: estructura, biosíntesis y función". Progreso en la investigación de lípidos . 44 (4): 235–258. doi :10.1016/j.plipres.2005.05.002. PMID 16019076.
- ^ ab Raetz CR, Garrett TA, Reynolds CM, Shaw WA, Moore JD, Smith DC, et al. (mayo de 2006). "Kdo2-Lipid A de Escherichia coli, una endotoxina definida que activa los macrófagos a través de TLR-4". Journal of Lipid Research . 47 (5): 1097–1111. doi : 10.1194/jlr.M600027-JLR200 . hdl : 10919/74310 . PMID 16479018.
- ^ Walsh CT (marzo de 2004). "Antibióticos policétidos y peptídicos no ribosomales: modularidad y versatilidad". Science . 303 (5665): 1805–1810. Bibcode :2004Sci...303.1805W. doi :10.1126/science.1094318. PMID 15031493. S2CID 44858908.
- ^ Caffrey P, Aparicio JF, Malpartida F, Zotchev SB (2008). "Ingeniería biosintética de macrólidos poliénicos hacia la generación de agentes antifúngicos y antiparasitarios mejorados". Temas actuales en química medicinal . 8 (8): 639–653. doi :10.2174/156802608784221479. hdl : 10197/8333 . PMID 18473889.
- ^ Minto RE, Blacklock BJ (julio de 2008). "Biosíntesis y función de poliacetilenos y productos naturales afines". Progreso en la investigación de lípidos . 47 (4): 233–306. doi :10.1016/j.plipres.2008.02.002. PMC 2515280. PMID 18387369 .
- ^ Coones RT, Green RJ, Frazier RA (julio de 2021). "Investigación de la composición del grupo de cabezas lipídicas dentro de las membranas epiteliales: una revisión sistemática". Soft Matter . 17 (28): 6773–6786. Bibcode :2021SMat...17.6773C. doi : 10.1039/D1SM00703C . ISSN 1744-683X. PMID 34212942. S2CID 235708094.
- ^ Heinz E. (1996). "Glicolípidos vegetales: estructura, aislamiento y análisis", págs. 211–332 en Advances in Lipid Methodology , vol. 3. WW Christie (ed.). Oily Press, Dundee. ISBN 978-0-9514171-6-4
- ^ Lyu, Jiabao; Gao, Renjun; Guo, Zheng (2021). "Galactosyldiacylglyceroles: De un aparato asociado a la fotosíntesis a un ensamblaje in vitro de estructura definida ". Revista de química agrícola y alimentaria . 69 (32): 8910–8928. doi :10.1021/acs.jafc.1c00204. PMID 33793221. S2CID 232761961.
- ^ Yashroy RC (1990). "Estudios de resonancia magnética de la organización dinámica de los lípidos en las membranas de los cloroplastos". Journal of Biosciences . 15 (4): 281–288. doi :10.1007/BF02702669. S2CID 360223.
- ^ van Meer G, Voelker DR, Feigenson GW (febrero de 2008). "Lípidos de membrana: dónde están y cómo se comportan". Nature Reviews Molecular Cell Biology . 9 (2): 112–124. doi :10.1038/nrm2330. PMC 2642958 . PMID 18216768.
- ^ Feigenson GW (noviembre de 2006). "Comportamiento de fases de mezclas de lípidos". Nature Chemical Biology . 2 (11): 560–563. doi :10.1038/nchembio1106-560. PMC 2685072 . PMID 17051225.
- ^ Wiggins PM (diciembre de 1990). "El papel del agua en algunos procesos biológicos". Microbiological Reviews . 54 (4): 432–449. doi :10.1128/MMBR.54.4.432-449.1990. PMC 372788 . PMID 2087221.
- ^ Raschke TM, Levitt M (mayo de 2005). "Los solutos no polares mejoran la estructura del agua dentro de las capas de hidratación al tiempo que reducen las interacciones entre ellas". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 102 (19): 6777–6782. doi : 10.1073/pnas.0500225102 . PMC 1100774 . PMID 15867152.
- ^ Segré D, Ben-Eli D, Deamer DW, Lancet D (2001). "El mundo de los lípidos" (PDF) . Orígenes de la vida y evolución de la biosfera . 31 (1–2): 119–145. Bibcode :2001OLEB...31..119S. doi :10.1023/A:1006746807104. PMID 11296516. S2CID 10959497. Archivado desde el original (PDF) el 11 de septiembre de 2008 . Consultado el 15 de marzo de 2015 .
- ^ Brasaemle DL (diciembre de 2007). "Serie de revisiones temáticas: biología de los adipocitos. La familia de proteínas estructurales de gotitas lipídicas perilipinas: estabilización de gotitas lipídicas y control de la lipólisis". Journal of Lipid Research . 48 (12): 2547–2559. doi : 10.1194/jlr.R700014-JLR200 . PMID 17878492.
- ^ Malinauskas T, Aricescu AR, Lu W, Siebold C, Jones EY (julio de 2011). "Mecanismo modular de inhibición de la señalización de Wnt por el factor inhibidor de Wnt 1". Nature Structural & Molecular Biology . 18 (8): 886–893. doi :10.1038/nsmb.2081. PMC 3430870 . PMID 21743455.
- ^ Malinauskas T (marzo de 2008). "Acoplamiento de ácidos grasos al dominio WIF del factor inhibidor de Wnt humano 1". Lípidos . 43 (3): 227–230. doi :10.1007/s11745-007-3144-3. PMID 18256869. S2CID 31357937.
- ^ Wang X (junio de 2004). "Señalización lipídica". Current Opinion in Plant Biology . 7 (3): 329–336. Bibcode :2004COPB....7..329W. doi :10.1016/j.pbi.2004.03.012. PMID 15134755.
- ^ Dinasarapu AR, Saunders B, Ozerlat I, Azam K, Subramaniam S (junio de 2011). "Páginas de moléculas de puerta de enlace de señalización: una perspectiva de modelo de datos". Bioinformática . 27 (12): 1736–1738. doi :10.1093/bioinformatics/btr190. PMC 3106186 . PMID 21505029.
- ^ Eyster KM (marzo de 2007). "La membrana y los lípidos como participantes integrales en la transducción de señales: transducción de señales lipídicas para el bioquímico no lipídico". Avances en la educación en fisiología . 31 (1): 5–16. doi :10.1152/advan.00088.2006. PMID 17327576. S2CID 9194419.
- ^ Hinkovska-Galcheva V, VanWay SM, Shanley TP, Kunkel RG (noviembre de 2008). "El papel de la esfingosina-1-fosfato y la ceramida-1-fosfato en la homeostasis del calcio". Current Opinion in Investigational Drugs . 9 (11): 1192–1205. PMID 18951299.
- ^ Saddoughi SA, Song P, Ogretmen B (2008). "Funciones de los esfingolípidos bioactivos en la biología y la terapéutica del cáncer". Lípidos en la salud y la enfermedad . Bioquímica subcelular. Vol. 49. págs. 413–440. doi :10.1007/978-1-4020-8831-5_16. ISBN . 978-1-4020-8830-8. PMC 2636716 . PMID 18751921.
- ^ Klein C, Malviya AN (enero de 2008). "Mecanismo de señalización del calcio nuclear por el inositol 1,4,5-trifosfato producido en el núcleo, la proteína quinasa C localizada en el núcleo y la proteína quinasa dependiente de AMP cíclico". Frontiers in Bioscience . 13 (13): 1206–1226. doi : 10.2741/2756 . PMID 17981624.
- ^ Boyce JA (agosto de 2008). "Eicosanoides en el asma, la inflamación alérgica y la defensa del huésped". Medicina molecular actual . 8 (5): 335–349. doi :10.2174/156652408785160989. PMID 18691060.
- ^ Bełtowski J (2008). "Receptores X del hígado (LXR) como dianas terapéuticas en la dislipidemia". Terapéutica cardiovascular . 26 (4): 297–316. doi : 10.1111/j.1755-5922.2008.00062.x . PMID 19035881.
- ^ Biermann M, Maueröder C, Brauner JM, Chaurio R, Janko C, Herrmann M, Muñoz LE (diciembre de 2013). "Código de superficie: señales biofísicas para la eliminación de células apoptóticas". Physical Biology . 10 (6): 065007. Bibcode :2013PhBio..10f5007B. doi :10.1088/1478-3975/10/6/065007. PMID 24305041. S2CID 23782770.
- ^ Indiveri C, Tonazzi A, Palmieri F (octubre de 1991). "Caracterización del transporte unidireccional de carnitina catalizado por el transportador de carnitina reconstituido a partir de mitocondrias de hígado de rata". Biochimica et Biophysica Acta (BBA) - Biomembranas . 1069 (1): 110–116. doi :10.1016/0005-2736(91)90110-t. PMID 1932043.
- ^ Parodi AJ, Leloir LF (abril de 1979). "El papel de los intermediarios lipídicos en la glicosilación de proteínas en la célula eucariota". Biochimica et Biophysica Acta (BBA) - Reseñas sobre biomembranas . 559 (1): 1–37. doi :10.1016/0304-4157(79)90006-6. PMID 375981.
- ^ Helenius A, Aebi M (marzo de 2001). "Funciones intracelulares de los glicanos ligados a N". Science . 291 (5512): 2364–2369. Bibcode :2001Sci...291.2364H. doi :10.1126/science.291.5512.2364. PMID 11269317. S2CID 7277949.
- ^ Nowicki M, Müller F, Frentzen M (abril de 2005). "Sintasa de cardiolipina de Arabidopsis thaliana". FEBS Letters . 579 (10): 2161–2165. Bibcode :2005FEBSL.579.2161N. doi : 10.1016/j.febslet.2005.03.007 . PMID 15811335. S2CID 21937549.
- ^ Gohil VM, Greenberg ML (febrero de 2009). "Biogénesis de la membrana mitocondrial: los fosfolípidos y las proteínas van de la mano". The Journal of Cell Biology . 184 (4): 469–472. doi :10.1083/jcb.200901127. PMC 2654137 . PMID 19237595.
- ^ Hoch FL (marzo de 1992). "Cardiolipinas y función de las biomembranas" (PDF) . Biochimica et Biophysica Acta (BBA) - Revisiones sobre biomembranas . 1113 (1): 71–133. doi :10.1016/0304-4157(92)90035-9. hdl : 2027.42/30145 . PMID 1550861.
- ^ "Esteroides". Elmhurst. edu . Archivado desde el original el 23 de octubre de 2011. Consultado el 10 de octubre de 2013 .
- ^ Chirala SS, Wakil SJ (noviembre de 2004). "Estructura y función de la sintetasa de ácidos grasos animales". Lípidos . 39 (11): 1045–1053. doi :10.1007/s11745-004-1329-9. PMID 15726818. S2CID 4043407.
- ^ White SW, Zheng J, Zhang YM (2005). "La biología estructural de la biosíntesis de ácidos grasos de tipo II". Revisión anual de bioquímica . 74 : 791–831. doi :10.1146/annurev.biochem.74.082803.133524. PMID 15952903.
- ^ Ohlrogge JB, Jaworski JG (junio de 1997). "Regulación de la síntesis de ácidos grasos". Revisión anual de fisiología vegetal y biología molecular de plantas . 48 : 109–136. doi :10.1146/annurev.arplant.48.1.109. PMID 15012259. S2CID 46348092.
- ^ Grochowski LL, Xu H, White RH (mayo de 2006). "Methanocaldococcus jannaschii utiliza una vía de mevalonato modificada para la biosíntesis de difosfato de isopentenilo". Journal of Bacteriology . 188 (9): 3192–3198. doi :10.1128/JB.188.9.3192-3198.2006. PMC 1447442 . PMID 16621811.
- ^ Lichtenthaler HK (junio de 1999). "La vía de la 1-didesoxi-D-xilulosa-5-fosfato en la biosíntesis de isoprenoides en plantas". Revisión anual de fisiología vegetal y biología molecular de plantas . 50 : 47–65. doi :10.1146/annurev.arplant.50.1.47. PMID 15012203.
- ^ ab Schroepfer GJ (1981). "Biosíntesis de esteroles". Revista anual de bioquímica . 50 : 585–621. doi :10.1146/annurev.bi.50.070181.003101. PMID 7023367.
- ^ Lees ND, Skaggs B, Kirsch DR, Bard M (marzo de 1995). "Clonación de los genes tardíos en la vía biosintética del ergosterol de Saccharomyces cerevisiae: una revisión". Lípidos . 30 (3): 221–226. doi :10.1007/BF02537824. PMID 7791529. S2CID 4019443.
- ^ ab Bhagavan NV (2002). Bioquímica médica. San Diego: Harcourt/Academic Press. ISBN 978-0-12-095440-7.
- ^ Russo GL (marzo de 2009). "Ácidos grasos poliinsaturados n-6 y n-3 en la dieta: de la bioquímica a las implicaciones clínicas en la prevención cardiovascular". Farmacología bioquímica . 77 (6): 937–946. doi :10.1016/j.bcp.2008.10.020. PMID 19022225.
- ^ Riediger ND, Othman RA, Suh M, Moghadasian MH (abril de 2009). "Una revisión sistemática de las funciones de los ácidos grasos n-3 en la salud y la enfermedad". Revista de la Asociación Dietética Estadounidense . 109 (4): 668–679. doi :10.1016/j.jada.2008.12.022. PMID 19328262.
- ^ Galli C, Risé P (2009). "Consumo de pescado, ácidos grasos omega 3 y enfermedad cardiovascular. La ciencia y los ensayos clínicos". Nutrición y salud . 20 (1): 11–20. doi :10.1177/026010600902000102. PMID 19326716. S2CID 20742062.
- ^ Micha R, Mozaffarian D (2008). "Ácidos grasos trans: efectos sobre la salud cardiometabólica e implicaciones para la política". Prostaglandinas, leucotrienos y ácidos grasos esenciales . 79 (3–5): 147–152. doi :10.1016/j.plefa.2008.09.008. PMC 2639783. PMID 18996687 .
- ^ Dalainas I, Ioannou HP (abril de 2008). "El papel de los ácidos grasos trans en la aterosclerosis, la enfermedad cardiovascular y el desarrollo infantil". Angiología internacional . 27 (2): 146–156. PMID 18427401.
- ^ Mozaffarian D, Willett WC (diciembre de 2007). "Ácidos grasos trans y riesgo cardiovascular: ¿una impronta cardiometabólica única?". Current Atherosclerosis Reports . 9 (6): 486–493. doi :10.1007/s11883-007-0065-9. PMID 18377789. S2CID 24998042.
- ^ Astrup A, Dyerberg J, Selleck M, Stender S (2008), "Transición nutricional y su relación con el desarrollo de la obesidad y enfermedades crónicas relacionadas", Obes Rev , 9 (S1): 48–52, doi :10.1111/j.1467-789X.2007.00438.x, PMID 18307699, S2CID 34030743
- ^ Astrup A (febrero de 2005). "El papel de la grasa dietética en la obesidad". Seminarios en Medicina Vascular . 5 (1): 40–47. doi :10.1055/s-2005-871740. PMID 15968579. S2CID 260372605.
- ^ Astrup A (2008). "Manejo dietético de la obesidad". Revista de nutrición parenteral y enteral . 32 (5): 575–577. doi :10.1177/0148607108321707. PMID 18753397.
- ^ Beresford SA, Johnson KC, Ritenbaugh C, Lasser NL, Snetselaar LG, Black HR, et al. (febrero de 2006). "Patrón dietético bajo en grasas y riesgo de cáncer colorrectal: ensayo controlado aleatorio de modificación dietética de la Women's Health Initiative". Revista de la Asociación Médica Estadounidense . 295 (6): 643–654. doi :10.1001/jama.295.6.643. PMID 16467233.
- ^ Howard BV, Manson JE, Stefanick ML, Beresford SA, Frank G, Jones B, Rodabough RJ, Snetselaar L, Thomson C, Tinker L, Vitolins M, Prentice R (enero de 2006). "Patrón dietético bajo en grasas y cambio de peso a lo largo de 7 años: el ensayo de modificación dietética de la Women's Health Initiative". Revista de la Asociación Médica Estadounidense . 295 (1): 39–49. doi :10.1001/jama.295.1.39. PMID 16391215.
- ^ "La fuente de nutrición". Escuela de Salud Pública TH Chan . Universidad de Harvard.
- ^ "Grasas y colesterol: ¡Fuera lo malo, dentro lo bueno! ¿Qué debería comer? - The Nutrition Source". Facultad de Salud Pública de Harvard.
Bibliografía
- Bhagavan NV (2002). Bioquímica médica. San Diego: Harcourt/Academic Press. ISBN 978-0-12-095440-7.
- Devlin TM (1997). Libro de texto de bioquímica: con correlaciones clínicas (4.ª ed.). Chichester: John Wiley & Sons. ISBN 978-0-471-17053-2.
- Stryer L, Berg JM, Tymoczko JL (2007). Bioquímica (6ª ed.). San Francisco: WH Freeman. ISBN 978-0-7167-8724-2.
- van Holde KE, Mathews CK (1996). Bioquímica (2.ª ed.). Menlo Park, California: Benjamin/Cummings Pub. Co. ISBN 978-0-8053-3931-4.
Enlaces externos
Introductorio
- Lista de sitios web relacionados con los lípidos
- Nature Lipidomics Gateway: resumen de las últimas investigaciones sobre lípidos
- Biblioteca de lípidos: referencia general sobre química y bioquímica de lípidos
- Cyberlipid.org – Recursos e historia de los lípidos.
- Simulaciones moleculares por computadora: modelado de membranas lipídicas
- Lípidos, membranas y tráfico de vesículas – La Biblioteca Virtual de Bioquímica, Biología Molecular y Biología Celular
Nomenclatura
- Nomenclatura IUPAC de lípidos
- Entrada del glosario de la IUPAC para la clase de moléculas lipídicas
Bases de datos
- MAPAS DE LÍPIDOS: bases de datos integrales de lípidos y genes/proteínas asociados a lípidos.
- LipidBank – Base de datos japonesa de lípidos y propiedades relacionadas, datos espectrales y referencias.
General
- ApolloLipids: proporciona información sobre la prevención y el tratamiento de la dislipidemia y las enfermedades cardiovasculares, así como programas de educación médica continua.
- Asociación Nacional de Lípidos: Organización de educación médica profesional para profesionales de la salud que buscan prevenir la morbilidad y la mortalidad derivadas de las dislipidemias y otros trastornos relacionados con el colesterol.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}