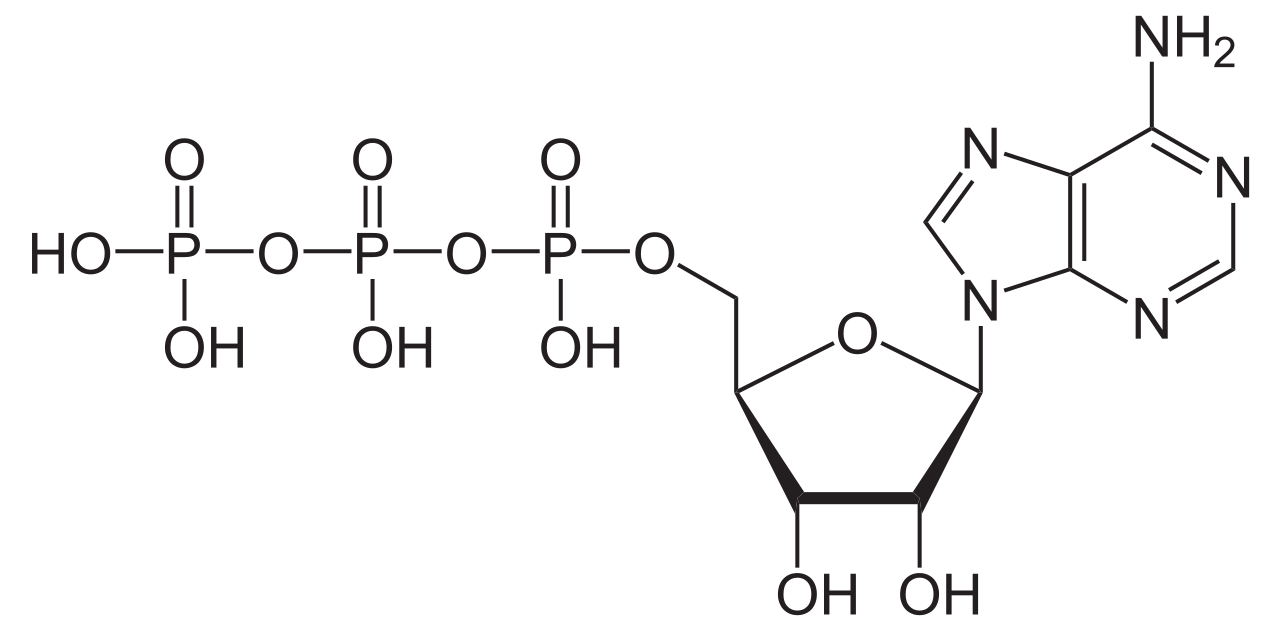
Trifosfato de adenosina
 | |||
| |||
| Nombres | |||
|---|---|---|---|
| Nombre IUPAC Adenosina 5′-(tetrahidrogenotrifosfato) | |||
| Nombre sistemático de la IUPAC Trifosfato de tetrahidrógeno de O 1 -{[(2 R ,3 S ,4 R ,5 R )-5-(6-amino-9 H -purin-9-il)-3,4-dihidroxioxolan-2-il]metil} | |||
| Identificadores | |||
| |||
Modelo 3D ( JSmol ) |
| ||
| EBICh |
| ||
| Química biológica |
| ||
| Araña química |
| ||
| Banco de medicamentos |
| ||
| Tarjeta informativa de la ECHA | 100.000.258 | ||
| |||
| BARRIL |
| ||
Identificador de centro de PubChem |
| ||
| UNIVERSIDAD |
| ||
Panel de control CompTox ( EPA ) |
| ||
| |||
| |||
| Propiedades | |||
| C10H16N5O13P3 | |||
| Masa molar | 507,18 g/mol | ||
| Densidad | 1,04 g/cm 3 (sal disódica) | ||
| Punto de fusión | 187 °C (369 °F; 460 K) sal disódica; se descompone | ||
| Acidez (p K a ) | 0,9, 1,4, 3,8, 6,5 | ||
| UV-vis (λmáx . ) | 259 nm [1] | ||
| Absorbancia | ε 259 = 15,4 mm −1 cm −1 [1] | ||
Salvo que se indique lo contrario, los datos se proporcionan para los materiales en su estado estándar (a 25 °C [77 °F], 100 kPa). | |||





El trifosfato de adenosina ( ATP ) es un trifosfato de nucleósido [2] que proporciona energía para impulsar y respaldar muchos procesos en las células vivas , como la contracción muscular , la propagación del impulso nervioso y la síntesis química . Presente en todas las formas de vida conocidas , a menudo se lo denomina la "unidad monetaria molecular " para la transferencia de energía intracelular . [3]
Cuando se consume en un proceso metabólico , el ATP se convierte en adenosín difosfato (ADP) o en adenosín monofosfato (AMP). Otros procesos regeneran el ATP. También es un precursor del ADN y el ARN , y se utiliza como coenzima . Un adulto humano promedio procesa alrededor de 50 kilogramos por día. [4]
Desde la perspectiva de la bioquímica , el ATP se clasifica como un nucleósido trifosfato , lo que indica que consta de tres componentes: una base nitrogenada ( adenina ), el azúcar ribosa y el trifosfato .
Estructura
El ATP consiste en una adenina unida por el átomo de nitrógeno #9 al átomo de carbono 1' de un azúcar ( ribosa ), que a su vez está unido en el átomo de carbono 5' del azúcar a un grupo trifosfato. En sus muchas reacciones relacionadas con el metabolismo, los grupos adenina y azúcar permanecen inalterados, pero el trifosfato se convierte en di- y monofosfato, dando lugar respectivamente a los derivados ADP y AMP . Los tres grupos fosforilo se etiquetan como alfa (α), beta (β) y, para el fosfato terminal, gamma (γ). [5]
En solución neutra, el ATP ionizado existe principalmente como ATP 4− , con una pequeña proporción de ATP 3− . [6]
Unión de cationes metálicos
El ATP es polianiónico y posee un grupo polifosfato potencialmente quelante , y se une a los cationes metálicos con alta afinidad. La constante de unión para el Mg2+
es ( 9 554 ). [7] La unión de un catión divalente , casi siempre magnesio , afecta fuertemente la interacción del ATP con varias proteínas. Debido a la fuerza de la interacción ATP-Mg 2+ , el ATP existe en la célula principalmente como un complejo con Mg2+
unido a los centros de oxígeno del fosfato. [6] [8]
Un segundo ion magnesio es fundamental para la unión del ATP en el dominio de la quinasa. [9] La presencia de Mg 2+ regula la actividad de la quinasa. [10] Es interesante desde la perspectiva del mundo del ARN que el ATP pueda transportar un ion Mg que cataliza la polimerización del ARN. [ cita requerida ]
Propiedades químicas
Las sales de ATP se pueden aislar como sólidos incoloros. [11]

El ATP es estable en soluciones acuosas entre pH 6,8 y 7,4 (en ausencia de catalizadores). A niveles de pH más extremos, se hidroliza rápidamente a ADP y fosfato. Las células vivas mantienen la relación de ATP a ADP en un punto a diez órdenes de magnitud del equilibrio, con concentraciones de ATP cinco veces superiores a la concentración de ADP. [12] [13] En el contexto de las reacciones bioquímicas, los enlaces POP se denominan con frecuencia enlaces de alta energía . [14]
Aspectos reactivos
La hidrólisis de ATP en ADP y fosfato inorgánico.
- ATP 4- (ac) + H 2 O (l) = ADP 3- (ac) + HPO 2- (ac) + H + (ac)
libera 20,5 kilojulios por mol (4,9 kcal/mol) de entalpía . Esto puede diferir en condiciones fisiológicas si el reactivo y los productos no están exactamente en estos estados de ionización. [15] Los valores de la energía libre liberada al escindir una unidad de fosfato (P i ) o de pirofosfato (PP i ) del ATP a concentraciones de estado estándar de 1 mol/L a pH 7 son: [16]
- ATP + H
2O → ADP + P i Δ G °' = −30,5 kJ/mol (−7,3 kcal/mol) - ATP + H
2O → AMP + PP i Δ G °' = −45,6 kJ/mol (−10,9 kcal/mol)
Estas ecuaciones abreviadas a un pH cercano a 7 se pueden escribir de forma más explícita (R = adenosilo ):
- [RO-P(O) 2 -OP(O) 2 -O-PO 3 ] 4− + H
2O → [RO-P(O) 2 -O-PO 3 ] 3− + [HPO 4 ] 2− + H + - [RO-P(O) 2 -OP(O) 2 -O-PO 3 ] 4− + H
2O → [RO-PO 3 ] 2− + [HO 3 P-O-PO 3 ] 3− + H +
En condiciones citoplasmáticas, donde la relación ADP/ATP está a 10 órdenes de magnitud del equilibrio, el Δ G es de alrededor de −57 kJ/mol. [12]
Junto con el pH, el cambio de energía libre de la hidrólisis de ATP también está asociado con la concentración de Mg 2+ , desde ΔG°' = −35,7 kJ/mol a una concentración de Mg 2+ de cero, hasta ΔG°' = −31 kJ/mol a [Mg 2+ ] = 5 mM. Concentraciones más altas de Mg 2+ disminuyen la energía libre liberada en la reacción debido a la unión de iones Mg 2+ a átomos de oxígeno de ATP con carga negativa a pH 7. [17]

Producción de AMP y ADP
Producción, condiciones aeróbicas
Una concentración intracelular típica de ATP puede ser de 1 a 10 μmol por gramo de tejido en una variedad de eucariotas. [18] La desfosforilación de ATP y la refosforilación de ADP y AMP ocurren repetidamente en el curso del metabolismo aeróbico. [19]
El ATP se puede producir mediante una serie de procesos celulares distintos; las tres vías principales en eucariotas son (1) la glucólisis , (2) el ciclo del ácido cítrico / fosforilación oxidativa y (3) la betaoxidación . El proceso general de oxidación de la glucosa a dióxido de carbono , la combinación de las vías 1 y 2, conocida como respiración celular , produce alrededor de 30 equivalentes de ATP a partir de cada molécula de glucosa. [20]
La producción de ATP por un eucariota aeróbico no fotosintético ocurre principalmente en las mitocondrias , que comprenden casi el 25% del volumen de una célula típica. [21]
Glucólisis
En la glucólisis, la glucosa y el glicerol se metabolizan a piruvato . La glucólisis genera dos equivalentes de ATP a través de la fosforilación del sustrato catalizada por dos enzimas, la fosfoglicerato quinasa (PGK) y la piruvato quinasa . También se producen dos equivalentes de nicotinamida adenina dinucleótido (NADH), que se pueden oxidar a través de la cadena de transporte de electrones y dar lugar a la generación de ATP adicional por la ATP sintasa . El piruvato generado como producto final de la glucólisis es un sustrato para el ciclo de Krebs . [22]
La glucólisis se considera que consta de dos fases con cinco pasos cada una. En la fase 1, "la fase preparatoria", la glucosa se convierte en 2 d-gliceraldehído-3-fosfato (g3p). Se invierte un ATP en el paso 1 y otro ATP en el paso 3. Los pasos 1 y 3 de la glucólisis se denominan "pasos de preparación". En la fase 2, dos equivalentes de g3p se convierten en dos piruvatos. En el paso 7, se producen dos ATP. Además, en el paso 10, se producen dos equivalentes más de ATP. En los pasos 7 y 10, se genera ATP a partir de ADP. Se forma una red de dos ATP en el ciclo de la glucólisis. La vía de la glucólisis se asocia más tarde con el ciclo del ácido cítrico, que produce equivalentes adicionales de ATP. [ cita requerida ]
Regulación
En la glucólisis, la hexoquinasa es inhibida directamente por su producto, la glucosa-6-fosfato, y la piruvato quinasa es inhibida por el propio ATP. El principal punto de control de la vía glucolítica es la fosfofructoquinasa (PFK), que es inhibida alostéricamente por altas concentraciones de ATP y activada por altas concentraciones de AMP. La inhibición de la PFK por el ATP es inusual ya que el ATP también es un sustrato en la reacción catalizada por la PFK; la forma activa de la enzima es un tetrámero que existe en dos conformaciones, de las cuales solo una se une al segundo sustrato, la fructosa-6-fosfato (F6P). La proteína tiene dos sitios de unión para el ATP: el sitio activo es accesible en cualquiera de las conformaciones de la proteína, pero la unión del ATP al sitio inhibidor estabiliza la conformación que se une mal al F6P. [22] Varias otras moléculas pequeñas pueden compensar el cambio inducido por ATP en la conformación de equilibrio y reactivar la PFK, incluyendo el AMP cíclico , los iones de amonio , el fosfato inorgánico y la fructosa-1,6- y -2,6-bifosfato. [22]
Ciclo del ácido cítrico
En la mitocondria , el piruvato es oxidado por el complejo piruvato deshidrogenasa al grupo acetilo , que se oxida completamente a dióxido de carbono por el ciclo del ácido cítrico (también conocido como el ciclo de Krebs ). Cada "vuelta" del ciclo del ácido cítrico produce dos moléculas de dióxido de carbono, un equivalente de ATP guanosina trifosfato (GTP) a través de la fosforilación a nivel de sustrato catalizada por la succinil-CoA sintetasa , ya que la succinil-CoA se convierte en succinato, tres equivalentes de NADH y un equivalente de FADH 2 . El NADH y el FADH 2 se reciclan (a NAD + y FAD , respectivamente) por fosforilación oxidativa , generando ATP adicional. La oxidación del NADH da como resultado la síntesis de 2-3 equivalentes de ATP, y la oxidación de un FADH 2 produce entre 1 y 2 equivalentes de ATP. [20] La mayoría del ATP celular se genera mediante este proceso. Aunque el ciclo del ácido cítrico en sí no involucra oxígeno molecular , es un proceso aeróbico obligado porque se utiliza O 2 para reciclar el NADH y el FADH 2 . En ausencia de oxígeno, el ciclo del ácido cítrico cesa. [21]
La generación de ATP por la mitocondria a partir del NADH citosólico depende de la lanzadera de malato-aspartato (y en menor medida, de la lanzadera de glicerol-fosfato ) porque la membrana mitocondrial interna es impermeable al NADH y al NAD + . En lugar de transferir el NADH generado, una enzima malato deshidrogenasa convierte el oxaloacetato en malato , que se transloca a la matriz mitocondrial. Otra reacción catalizada por la malato deshidrogenasa ocurre en la dirección opuesta, produciendo oxaloacetato y NADH a partir del malato recién transportado y del depósito interior de NAD + de la mitocondria . Una transaminasa convierte el oxaloacetato en aspartato para transportarlo de regreso a través de la membrana y hacia el espacio intermembrana. [21]
En la fosforilación oxidativa, el paso de electrones desde el NADH y el FADH2 a través de la cadena de transporte de electrones libera la energía necesaria para bombear protones fuera de la matriz mitocondrial y hacia el espacio intermembrana. Este bombeo genera una fuerza motriz protónica que es el efecto neto de un gradiente de pH y un gradiente de potencial eléctrico a través de la membrana mitocondrial interna. El flujo de protones a lo largo de este gradiente de potencial (es decir, desde el espacio intermembrana hacia la matriz) produce ATP por acción de la ATP sintasa. [23] Se producen tres ATP por vuelta.
Aunque el consumo de oxígeno parece fundamental para el mantenimiento de la fuerza motriz de protones, en caso de escasez de oxígeno ( hipoxia ), la acidosis intracelular (mediada por tasas glucolíticas mejoradas e hidrólisis de ATP ), contribuye al potencial de membrana mitocondrial e impulsa directamente la síntesis de ATP. [24]
La mayor parte del ATP sintetizado en las mitocondrias se utilizará para procesos celulares en el citosol; por lo tanto, debe exportarse desde su sitio de síntesis en la matriz mitocondrial. El movimiento de salida del ATP se ve favorecido por el potencial electroquímico de la membrana porque el citosol tiene una carga relativamente positiva en comparación con la matriz relativamente negativa. Por cada ATP transportado, cuesta 1 H + . Producir un ATP cuesta alrededor de 3 H + . Por lo tanto, fabricar y exportar un ATP requiere 4H + . La membrana interna contiene un antiportador , la translocasa ADP/ATP, que es una proteína integral de membrana que se utiliza para intercambiar ATP recién sintetizado en la matriz por ADP en el espacio intermembrana. [25]
Regulación
El ciclo del ácido cítrico está regulado principalmente por la disponibilidad de sustratos clave, en particular la relación de NAD + a NADH y las concentraciones de calcio , fosfato inorgánico, ATP, ADP y AMP. El citrato , el ion que da nombre al ciclo, es un inhibidor de retroalimentación de la citrato sintasa y también inhibe la PFK, lo que proporciona un vínculo directo entre la regulación del ciclo del ácido cítrico y la glucólisis. [22]
Beta oxidación
En presencia de aire y varios cofactores y enzimas, los ácidos grasos se convierten en acetil-CoA . La vía se llama beta-oxidación . Cada ciclo de beta-oxidación acorta la cadena de ácidos grasos en dos átomos de carbono y produce un equivalente de cada uno de acetil-CoA, NADH y FADH 2 . El acetil-CoA es metabolizado por el ciclo del ácido cítrico para generar ATP, mientras que el NADH y el FADH 2 son utilizados por fosforilación oxidativa para generar ATP. Se generan docenas de equivalentes de ATP por la beta-oxidación de una única cadena larga de acilo. [26]
Regulación
En la fosforilación oxidativa, el punto de control clave es la reacción catalizada por la citocromo c oxidasa , que está regulada por la disponibilidad de su sustrato, la forma reducida del citocromo c . La cantidad de citocromo c reducido disponible está directamente relacionada con las cantidades de otros sustratos:

lo que implica directamente esta ecuación:
![{\displaystyle {\frac {[\mathrm {cyt~c_{red}} ]}{[\mathrm {cyt~c_{ox}} ]}}=\left({\frac {[\mathrm {NADH} ] }{[\mathrm {NAD} ]^{+}}}\right)^{\frac {1}{2}}\left({\frac {[\mathrm {ADP} ][\mathrm {P_{i) }} ]}{[\mathrm {ATP} ]}}\right)K_{\mathrm {eq} }}](https://wikimedia.org/api/rest_v1/media/math/render/svg/baed008d658c507750599ae155a9a1c11da873e7)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Por lo tanto, una alta relación de [NADH] a [NAD + ] o una alta relación de [ADP] [P i ] a [ATP] implican una alta cantidad de citocromo c reducido y un alto nivel de actividad de la citocromo c oxidasa. [22] Un nivel adicional de regulación es introducido por las tasas de transporte de ATP y NADH entre la matriz mitocondrial y el citoplasma. [25]
Cetosis
Los cuerpos cetónicos pueden utilizarse como combustibles, produciendo 22 moléculas de ATP y 2 de GTP por molécula de acetoacetato cuando se oxidan en las mitocondrias. Los cuerpos cetónicos se transportan desde el hígado a otros tejidos, donde el acetoacetato y el beta -hidroxibutirato pueden reconvertirse en acetil-CoA para producir equivalentes reductores (NADH y FADH 2 ), a través del ciclo del ácido cítrico. Los cuerpos cetónicos no pueden ser utilizados como combustible por el hígado, porque el hígado carece de la enzima β-cetoacil-CoA transferasa, también llamada tiolasa . El acetoacetato en bajas concentraciones es absorbido por el hígado y sufre una desintoxicación a través de la vía del metilglioxal que termina con el lactato. El acetoacetato en altas concentraciones es absorbido por células distintas de las del hígado y entra en una vía diferente a través del 1,2-propanodiol . Aunque la vía sigue una serie diferente de pasos que requieren ATP, el 1,2-propanodiol puede convertirse en piruvato. [27]
Producción, condiciones anaeróbicas
La fermentación es el metabolismo de compuestos orgánicos en ausencia de aire. Implica la fosforilación a nivel de sustrato en ausencia de una cadena de transporte de electrones respiratoria . La ecuación para la reacción de la glucosa para formar ácido láctico es:
- do
6yo
12Oh
6+ 2 ADP + 2 Pi → 2 CH
3CH(OH)COOH + 2 ATP + 2 H
2Oh
La respiración anaeróbica es la respiración en ausencia de O
2Los procariotas pueden utilizar una variedad de aceptores de electrones, entre ellos el nitrato , el sulfato y el dióxido de carbono.
Reposición de ATP por nucleósido difosfato quinasas
El ATP también se puede sintetizar a través de varias reacciones llamadas de "reposición" catalizadas por las familias de enzimas de las nucleósido difosfato quinasas (NDK), que utilizan otros nucleósidos trifosfatos como donadores de fosfato de alta energía, y la familia ATP:guanido-fosfotransferasa . [ cita requerida ]
Producción de ATP durante la fotosíntesis
En las plantas, el ATP se sintetiza en la membrana tilacoide del cloroplasto . El proceso se denomina fotofosforilación . La "maquinaria" es similar a la de las mitocondrias, excepto que se utiliza la energía de la luz para bombear protones a través de una membrana para producir una fuerza protón-motriz. La ATP sintasa se activa exactamente como en la fosforilación oxidativa. [28] Parte del ATP producido en los cloroplastos se consume en el ciclo de Calvin , que produce azúcares triosa .
Reciclaje de ATP
La cantidad total de ATP en el cuerpo humano es de aproximadamente 0,1 mol/L . [29] La mayor parte del ATP se recicla a partir del ADP mediante los procesos antes mencionados. Por lo tanto, en cualquier momento dado, la cantidad total de ATP + ADP permanece bastante constante.
La energía utilizada por las células humanas en un adulto requiere la hidrólisis de 100 a 150 mol/L de ATP diariamente, lo que significa que un humano normalmente utilizará el equivalente a su peso corporal en ATP a lo largo del día. [30] Cada equivalente de ATP se recicla entre 1000 y 1500 veces durante un solo día ( 150 / 0,1 = 1500 ), [29] a aproximadamente 9×10 20 moléculas/s. [29]
{kind=link}

Funciones bioquímicas
Señalización intracelular
El ATP participa en la transducción de señales al actuar como sustrato para las quinasas, enzimas que transfieren grupos fosfato. Las quinasas son las proteínas de unión al ATP más comunes. Comparten una pequeña cantidad de pliegues comunes. [31] La fosforilación de una proteína por una quinasa puede activar una cascada como la cascada de la proteína quinasa activada por mitógeno . [32]
El ATP también es un sustrato de la adenilato ciclasa , más comúnmente en las vías de transducción de señales del receptor acoplado a proteína G y se transforma en un segundo mensajero , el AMP cíclico, que participa en la activación de señales de calcio mediante la liberación de calcio de los depósitos intracelulares. [33] Esta forma de transducción de señales es particularmente importante en la función cerebral, aunque está involucrada en la regulación de una multitud de otros procesos celulares. [34]
Síntesis de ADN y ARN
El ATP es uno de los cuatro monómeros necesarios para la síntesis de ARN . El proceso es promovido por las ARN polimerasas . [35] Un proceso similar ocurre en la formación de ADN, excepto que el ATP se convierte primero en el desoxirribonucleótido dATP. Al igual que muchas reacciones de condensación en la naturaleza, la replicación y la transcripción del ADN también consumen ATP.
Activación de aminoácidos en la síntesis de proteínas
Las enzimas aminoacil-ARNt sintetasa consumen ATP en la unión del ARNt a los aminoácidos, formando complejos aminoacil-ARNt. La aminoacil transferasa une el aminoácido AMP al ARNt. La reacción de acoplamiento se desarrolla en dos pasos:
- aa + ATP ⟶ aa-AMP + PP i
- aa-AMP + ARNt ⟶ aa-ARNt + AMP
El aminoácido está acoplado al penúltimo nucleótido en el extremo 3' del ARNt (la A en la secuencia CCA) a través de un enlace éster (desplazarse por encima en la ilustración).
Transportador de casete de unión de ATP
El transporte de sustancias químicas fuera de una célula en contra de un gradiente suele estar asociado a la hidrólisis de ATP. El transporte está mediado por transportadores de casete de unión de ATP . El genoma humano codifica 48 transportadores ABC, que se utilizan para exportar fármacos, lípidos y otros compuestos. [36]
Señalización extracelular y neurotransmisión
Las células secretan ATP para comunicarse con otras células en un proceso llamado señalización purinérgica . El ATP sirve como neurotransmisor en muchas partes del sistema nervioso, modula el movimiento ciliar, afecta el suministro de oxígeno vascular, etc. El ATP se secreta directamente a través de la membrana celular mediante proteínas de canal [37] [38] o se bombea a vesículas [39] que luego se fusionan con la membrana. Las células detectan ATP utilizando las proteínas receptoras purinérgicas P2X y P2Y [40] . Se ha demostrado que el ATP es una molécula de señalización de importancia crítica para las interacciones entre la microglía y las neuronas en el cerebro adulto [41] , así como durante el desarrollo cerebral [42] . Además, la señalización de ATP inducida por la lesión tisular es un factor importante en los cambios rápidos del fenotipo de la microglía [43] .
Contracción muscular
El ATP alimenta las contracciones musculares . [44] Las contracciones musculares están reguladas por vías de señalización, aunque los diferentes tipos de músculos están regulados por vías y estímulos específicos en función de su función particular. Sin embargo, en todos los tipos de músculos, la contracción la realizan las proteínas actina y miosina . [45]
El ATP se une inicialmente a la miosina. Cuando la ATPasa hidroliza el ATP unido en ADP y fosfato inorgánico , la miosina se posiciona de manera que pueda unirse a la actina. La miosina unida por ADP y P i forma puentes cruzados con la actina y la posterior liberación de ADP y P i libera energía como golpe de potencia. El golpe de potencia hace que el filamento de actina se deslice más allá del filamento de miosina, acortando el músculo y provocando una contracción. Otra molécula de ATP puede entonces unirse a la miosina, liberándola de la actina y permitiendo que este proceso se repita. [45] [46]
Solubilidad de proteínas
Recientemente se ha propuesto que el ATP actúa como un hidrótropo biológico [47] y se ha demostrado que afecta la solubilidad de todo el proteoma. [48]
Orígenes abiogénicos
El acetilfosfato (AcP), un precursor del ATP, puede sintetizarse fácilmente con rendimientos modestos a partir de tioacetato a pH 7 y 20 °C y pH 8 y 50 °C, aunque el acetilfosfato es menos estable en temperaturas más cálidas y condiciones alcalinas que en condiciones más frías y ácidas a neutras. No puede promover la polimerización de ribonucleótidos y aminoácidos y solo fue capaz de fosforilar compuestos orgánicos. Se demostró que puede promover la agregación y estabilización de AMP en presencia de Na + , la agregación de nucleótidos podría promover la polimerización por encima de 75 °C en ausencia de Na + . Es posible que la polimerización promovida por AcP pueda ocurrir en superficies minerales. [49] Se demostró que el ADP solo puede ser fosforilado a ATP por AcP y otros trifosfatos de nucleósidos no fueron fosforilados por AcP. Esto podría explicar por qué todas las formas de vida usan ATP para impulsar reacciones bioquímicas. [50]
Análogos del ATP
Los laboratorios de bioquímica suelen utilizar estudios in vitro para explorar los procesos moleculares dependientes del ATP. Los análogos del ATP también se utilizan en la cristalografía de rayos X para determinar la estructura de una proteína en complejo con el ATP, a menudo junto con otros sustratos. [ cita requerida ]
Se necesitan inhibidores enzimáticos de enzimas dependientes de ATP, como las quinasas, para examinar los sitios de unión y los estados de transición involucrados en las reacciones dependientes de ATP. [ cita requerida ]
La mayoría de los análogos de ATP útiles no se pueden hidrolizar como lo haría el ATP; en cambio, atrapan la enzima en una estructura estrechamente relacionada con el estado unido al ATP. El adenosín 5'-(γ-tiotrifosfato) es un análogo de ATP extremadamente común en el que uno de los oxígenos del gamma-fosfato es reemplazado por un átomo de azufre ; este anión se hidroliza a una velocidad drásticamente más lenta que el propio ATP y funciona como un inhibidor de los procesos dependientes del ATP. En los estudios cristalográficos, los estados de transición de hidrólisis se modelan mediante el ion vanadato unido.
Se recomienda tener precaución al interpretar los resultados de experimentos que utilizan análogos de ATP, ya que algunas enzimas pueden hidrolizarlos a tasas apreciables en altas concentraciones. [51]
Uso médico
El ATP se utiliza por vía intravenosa para algunas enfermedades relacionadas con el corazón. [52]
Historia
El ATP fue descubierto en 1929 por Karl Lohmann [53] y Jendrassik [54] e, independientemente, por Cyrus Fiske y Yellapragada Subba Rao de la Facultad de Medicina de Harvard , [55] ambos equipos compitiendo entre sí para encontrar un ensayo para el fósforo.
Fritz Albert Lipmann lo propuso en 1941 como intermediario entre las reacciones que producen energía y las que la requieren en las células. [56]
Fue sintetizado por primera vez en el laboratorio por Alexander Todd en 1948, [57] y recibió el Premio Nobel de Química en 1957 en parte por este trabajo.
El Premio Nobel de Química de 1978 fue otorgado a Peter Dennis Mitchell por el descubrimiento del mecanismo quimiosmótico de la síntesis de ATP.
El Premio Nobel de Química de 1997 se dividió, la mitad conjuntamente para Paul D. Boyer y John E. Walker "por su elucidación del mecanismo enzimático subyacente a la síntesis de trifosfato de adenosina (ATP)" y la otra mitad para Jens C. Skou "por el primer descubrimiento de una enzima transportadora de iones, Na + , K + -ATPasa". [58]
Véase también
Referencias
- ^ ab "Información del producto de la sal disódica de adenosina 5'-trifosfato" (PDF) . Sigma. Archivado (PDF) del original el 23 de marzo de 2019 . Consultado el 22 de marzo de 2019 .
- ^ Dunn, Jacob; Grider, Michael H. (2023). "Fisiología, trifosfato de adenosina". StatPearls . StatPearls Publishing. PMID 31985968 . Consultado el 13 de noviembre de 2023 .
- ^ Knowles, J. R. (1980). "Reacciones de transferencia de fosforilo catalizadas por enzimas". Annu. Rev. Biochem . 49 : 877–919. doi :10.1146/annurev.bi.49.070180.004305. PMID 6250450.
- ^ "Un individuo promedio con una dieta diaria de 8000 kJ y una eficiencia del 30% en la conversión de alimentos en energía química sintetizará (e hidrolizará) alrededor de 50 kg de ATP durante 1 día". Wilkens, Stephan (2008). "Síntesis de ATP, química de". Wiley Encyclopedia of Chemical Biology . doi :10.1002/9780470048672.wecb648. ISBN 9780471754770.
- ^ Dunn, Jacob; Grider, Michael H. (2023), "Fisiología, trifosfato de adenosina", StatPearls , Treasure Island (FL): StatPearls Publishing, PMID 31985968 , consultado el 28 de septiembre de 2023
- ^ ab Storer, A.; Cornish-Bowden, A. (1976). "Concentración de MgATP2− y otros iones en solución. Cálculo de las concentraciones reales de especies presentes en mezclas de iones asociados". Biochem. J . 159 (1): 1–5. doi :10.1042/bj1590001. PMC 1164030 . PMID 11772.
- ^ Wilson, J.; Chin, A. (1991). "Quelación de cationes divalentes por ATP, estudiada por calorimetría de titulación". Anal. Biochem . 193 (1): 16–19. doi :10.1016/0003-2697(91)90036-S. PMID 1645933.
- ^ Garfinkel, L.; Altschuld, R.; Garfinkel, D. (1986). "Magnesio en el metabolismo energético cardíaco". J. Mol. Cell. Cardiol . 18 (10): 1003–1013. doi :10.1016/S0022-2828(86)80289-9. PMID 3537318.
- ^ Saylor, P.; Wang, C.; Hirai, T.; Adams, J. (1998). "Un segundo ion magnesio es fundamental para la unión de ATP en el dominio quinasa de la oncoproteína v-Fps". Bioquímica . 37 (36): 12624–12630. doi :10.1021/bi9812672. PMID 9730835.
- ^ Lin, X.; Ayrapetov, M; Sun, G. (2005). "Caracterización de las interacciones entre el sitio activo de una proteína tirosina quinasa y un activador de metal divalente". BMC Biochem . 6 : 25. doi : 10.1186/1471-2091-6-25 . PMC 1316873 . PMID 16305747.
- ^ Budavari, Susan, ed. (2001), El índice Merck: una enciclopedia de productos químicos, fármacos y productos biológicos (13.ª ed.), Merck, ISBN 0911910131
- ^ ab Ferguson, S. J.; Nicholls, David; Ferguson, Stuart (2002). Bioenergética 3 (3.ª ed.). San Diego, CA: Academic. ISBN 978-0-12-518121-1.
- ^ Berg, JM; Tymoczko, JL; Stryer, L. (2003). Bioquímica . Nueva York, Nueva York: W. H. Freeman. pag. 376.ISBN 978-0-7167-4684-3.
- ^ Chance, B.; Lees, H.; Postgate, J. G. (1972). "El significado de "flujo de electrones invertido" y "electrón de alta energía" en bioquímica". Nature . 238 (5363): 330–331. Bibcode :1972Natur.238..330C. doi :10.1038/238330a0. PMID 4561837. S2CID 4298762.
- ^ Gajewski, E.; Steckler, D.; Goldberg, R. (1986). "Termodinámica de la hidrólisis de adenosina 5'-trifosfato a adenosina 5'-difosfato". J. Biol. Chem . 261 (27): 12733–12737. doi : 10.1016/S0021-9258(18)67153-4 . PMID: 3528161.
- ^ Berg, Jeremy M.; Tymoczko, John L.; Stryer, Lubert (2007). Bioquímica (6ª ed.). Nueva York, Nueva York: W. H. Freeman. pag. 413.ISBN 978-0-7167-8724-2.
- ^ Garrett, Reginald H.; Grisham, Charles M. (2016). Bioquímica (6.ª ed.). Cengage Learning. pág. 68. ISBN 978-1305577206.
- ^ Beis, I.; Newsholme, E. A. (1 de octubre de 1975). "El contenido de nucleótidos de adenina, fosfágenos y algunos intermediarios glucolíticos en músculos en reposo de vertebrados e invertebrados". Biochem. J. 152 ( 1): 23–32. doi :10.1042/bj1520023. PMC 1172435. PMID 1212224 .
- ^ "Trifosfato de adenosina". Britannica. 11 de noviembre de 2023. Consultado el 1 de diciembre de 2023 .
- ^ ab Rich, P. R. (2003). "La maquinaria molecular de la cadena respiratoria de Keilin". Biochem. Soc. Trans . 31 (6): 1095–1105. doi :10.1042/BST0311095. PMID 14641005.
- ^ abc Lodish, H.; Berk, A.; Matsudaira, P.; Kaiser, C. A.; Krieger, M.; Scott, M. P.; Zipursky, S. L.; Darnell, J. (2004). Biología celular molecular (5.ª ed.). Nueva York, NY: W. H. Freeman. ISBN 978-0-7167-4366-8.
- ^ abcdeVoet, D.; Voet, JG (2004). Bioquímica . vol. 1 (3ª ed.). Hoboken, Nueva Jersey: Wiley. ISBN 978-0-471-19350-0.
- ^ Abrahams, J.; Leslie, A.; Lutter, R.; Walker, J. (1994). "Estructura con una resolución de 2,8 Å de la F1-ATPasa de mitocondrias de corazón bovino". Nature . 370 (6491): 621–628. Bibcode :1994Natur.370..621A. doi :10.1038/370621a0. PMID 8065448. S2CID 4275221.
- ^ Devaux, JBL; Hedges, CP; Hickey, AJR (enero de 2019). "La acidosis mantiene la función de las mitocondrias cerebrales en peces de triple aleta tolerantes a la hipoxia: ¿una estrategia para sobrevivir a la exposición hipóxica aguda?". Front Physiol . 9, 1914: 1941. doi : 10.3389/fphys.2018.01941 . PMC 6346031 . PMID 30713504.
- ^ ab Dahout-Gonzalez, C.; Nury, H.; Trézéguet, V.; Lauquin, G.; Pebay-Peyroula, E.; Brandolin, G. (2006). "Aspectos moleculares, funcionales y patológicos del transportador mitocondrial de ADP/ATP". Fisiología . 21 (4): 242–249. doi :10.1152/physiol.00005.2006. PMID 16868313.
- ^ Ronnett, G.; Kim, E.; Landree, L.; Tu, Y. (2005). "Metabolismo de ácidos grasos como objetivo para el tratamiento de la obesidad". Physiol. Behav . 85 (1): 25–35. doi :10.1016/j.physbeh.2005.04.014. PMID 15878185. S2CID 24865576.
- ^ "Sistema Integrado de Información de Riesgos" (PDF) . 2013-03-15. Archivado (PDF) desde el original el 2015-09-24 . Consultado el 2019-02-01 .
- ^ Allen, J. (2002). "Fotosíntesis de electrones ATP, bombas de protones, rotores y equilibrio". Cell . 110 (3): 273–276. doi : 10.1016/S0092-8674(02)00870-X . PMID 12176312. S2CID 1754660.
- ^ abc Fuhrman, Bradley P.; Zimmerman, Jerry J. (2011). Cuidados intensivos pediátricos. Elsevier. pág. 1061. ISBN 978-0-323-07307-3. Recuperado el 16 de mayo de 2020 .
- ^ Fuhrman, Bradley P.; Zimmerman, Jerry J. (2011). Cuidados intensivos pediátricos. Elsevier. págs. 1058–1072. ISBN 978-0-323-07307-3. Recuperado el 16 de mayo de 2020 .
- ^ Scheeff, E.; Bourne, P. (2005). "Evolución estructural de la superfamilia de proteínas similares a las quinasas". PLOS Comput. Biol . 1 (5): e49. Bibcode :2005PLSCB...1...49S. doi : 10.1371/journal.pcbi.0010049 . PMC 1261164 . PMID 16244704.
- ^ Mishra, N.; Tuteja, R.; Tuteja, N. (2006). "Señalización a través de redes de quinasas MAP en plantas". Arch. Biochem. Biophys . 452 (1): 55–68. doi :10.1016/j.abb.2006.05.001. PMID 16806044.
- ^ Kamenetsky, M.; Middelhaufe, S.; Bank, E.; Levin, L.; Buck, J.; Steegborn, C. (2006). "Detalles moleculares de la generación de AMPc en células de mamíferos: una historia de dos sistemas". J. Mol. Biol . 362 (4): 623–639. doi :10.1016/j.jmb.2006.07.045. PMC 3662476. PMID 16934836 .
- ^ Hanoune, J.; Defer, N. (2001). "Regulación y función de las isoformas de la adenilil ciclasa". Annu. Rev. Pharmacol. Toxicol . 41 (1): 145–174. doi :10.1146/annurev.pharmtox.41.1.145. PMID 11264454.
- ^ Joyce, C. M.; Steitz, T. A. (1995). "Estructuras y funciones de la polimerasa: ¿variaciones sobre un tema?". J. Bacteriol . 177 (22): 6321–6329. doi : 10.1128/jb.177.22.6321-6329.1995. PMC 177480. PMID 7592405.
- ^ Borst, P.; Elferink, R. Oude (2002). "Transportadores ABC de mamíferos en la salud y la enfermedad" (PDF) . Revisión anual de bioquímica . 71 : 537–592. doi :10.1146/annurev.biochem.71.102301.093055. PMID 12045106. S2CID 34707074. Archivado (PDF) desde el original el 21 de abril de 2018 . Consultado el 20 de abril de 2018 .
- ^ Romanov, romano A.; Lasher, Robert S.; Alto, Brigit; Savidge, Logan E.; Lawson, Adán; Rogachevskaja, Olga A.; Zhao, haitiano; Rogachevski, Vadim V.; Bystrova, Marina F.; Churbanov, Gleb D.; Adameyko, Igor; Harkány, Tibor; Yang, Ruibiao; Kidd, Grahame J.; Marambaud, Philippe; Kinnamon, John C.; Kolesnikov, Stanislav S.; Dedo, Thomas E. (2018). "Sinapsis químicas sin vesículas sinápticas: neurotransmisión purinérgica a través de un complejo de señalización mitocondrial-canal CALHM1". Señalización científica . 11 (529): eaao1815. doi : 10.1126/scisignal.aao1815. ISSN 1945-0877. Número de modelo : PMID 29739879.
- ^ Dahl, Gerhard (2015). "Liberación de ATP a través de los canales pannexones". Philosophical Transactions of the Royal Society B: Biological Sciences . 370 (1672): 20140191. doi :10.1098/rstb.2014.0191. ISSN 0962-8436. PMC 4455760 . PMID 26009770.
- ^ Larsson, Max; Sawada, Keisuke; Morland, Cecilie; Hiasa, Miki; Ormel, Lasse; Moriyama, Yoshinori; Gundersen, Vidar (2012). "Identificación funcional y anatómica de un transportador vesicular que media la liberación neuronal de ATP". Corteza cerebral . 22 (5): 1203–1214. doi : 10.1093/cercor/bhr203 . ISSN 1460-2199. PMID 21810784.
- ^ Puchałowicz, Kamila; Tarnowski, Maciej; Baranowska-Bosiacka, Irena; Chlubek, Dariusz; Dziedziejko, Violetta (18 de diciembre de 2014). "Receptores P2X y P2Y: papel en la fisiopatología del sistema nervioso". Revista Internacional de Ciencias Moleculares . 15 (12): 23672–23704. doi : 10.3390/ijms151223672 . ISSN 1422-0067. PMC 4284787 . PMID 25530618.
- ^ Csaba, Cserep; Balazs, Pósfai (2020). "La microglía controla y protege la función neuronal a través de uniones purinérgicas somáticas especializadas". Science . doi :10.1126/science.aax6752. PMID 31831638.
- ^ Csaba, Cserep; Anett, Schwarcz D (2022). "Control microglial del desarrollo neuronal a través de uniones purinérgicas somáticas". Cell Reports . doi :10.1016/j.celrep.2022.111369. PMID 36130488.
- ^ Peter, Berki; Csaba, Cserep; Zsuzsanna, Környei (2024). "La microglía contribuye a la sincronía neuronal a pesar de la transformación fenotípica relacionada con ATP endógeno en cortes agudos de cerebro de ratón". Nature Communications . doi :10.1038/s41467-024-49773-1. PMC 11208608 . PMID 38926390.
- ^ Hultman, E.; Greenhaff, PL (1991). "Metabolismo energético del músculo esquelético y fatiga durante el ejercicio intenso en el hombre". Science Progress . 75 (298 Pt 3-4): 361–370. ISSN 0036-8504. PMID 1842855.
- ^ ab Kuo, Ivana Y.; Ehrlich, Barbara E. (febrero de 2015). "Señalización en la contracción muscular". Cold Spring Harbor Perspectives in Biology . 7 (2): a006023. doi :10.1101/cshperspect.a006023. ISSN 1943-0264. PMC 4315934 . PMID 25646377.
- ^ "38.17: Contracción muscular y locomoción - ATP y contracción muscular". Biology LibreTexts . 2018-07-16 . Consultado el 2024-05-01 .
- ^ Hyman, Anthony A.; Krishnan, Yamuna; Alberti, Simon; Wang, Jie; Saha, Shambaditya; Malinovska, Liliana; Patel, Avinash (19 de mayo de 2017). "ATP como hidrótropo biológico". Science . 356 (6339): 753–756. Bibcode :2017Sci...356..753P. doi :10.1126/science.aaf6846. ISSN 0036-8075. PMID 28522535. S2CID 24622983.
- ^ Savitski, Mikhail M.; Bantscheff, Marcus; Huber, Wolfgang; Dominic Helm; Günthner, Ina; Werner, Thilo; Kurzawa, Nils; Sridharan, Sindhuja (11 de marzo de 2019). "El perfil de solubilidad y estabilidad térmica de todo el proteoma revela funciones reguladoras distintas para el ATP". Nature Communications . 10 (1): 1155. Bibcode :2019NatCo..10.1155S. doi :10.1038/s41467-019-09107-y. ISSN 2041-1723. PMC 6411743 . PMID 30858367.
- ^ Whicher, Alexandra; Camprubi, Eloi; Pinna, Silvana; Herschy, Barry; Lane, Nick (1 de junio de 2018). "El fosfato de acetilo como moneda de energía primordial en el origen de la vida". Orígenes de la vida y evolución de las biosferas . 48 (2): 159–179. Bibcode :2018OLEB...48..159W. doi :10.1007/s11084-018-9555-8. ISSN 1573-0875. PMC 6061221 . PMID 29502283.
- ^ "La química antigua puede explicar por qué los seres vivos utilizan el ATP como la moneda energética universal: un paso temprano en la evolución metabólica preparó el terreno para el surgimiento del ATP como portador de energía universal". ScienceDaily . Consultado el 27 de agosto de 2023 .
- ^ Resetar, A. M.; Chalovich, J. M. (1995). "Adenosina 5′-(gamma-tiotrifosfato): un análogo de ATP que debe utilizarse con precaución en estudios de contracción muscular". Bioquímica . 34 (49): 16039–16045. doi :10.1021/bi00049a018. PMID 8519760.
- ^ Pelleg, Amir; Kutalek, Steven P.; Flammang, Daniel; Benditt, David (febrero de 2012). "ATPace: adenosina 5′-trifosfato inyectable". Señalización purinérgica . 8 (Supl 1): 57–60. doi :10.1007/s11302-011-9268-1. ISSN 1573-9538. PMC 3265710 . PMID 22057692.
- ^ Lohmann, K. (agosto de 1929). "Über die Pyrophosphatfraktion im Muskel" [Sobre la fracción de pirofosfato en el músculo]. Naturwissenschaften (en alemán). 17 (31): 624–625. Código bibliográfico : 1929NW.....17..624.. doi : 10.1007/BF01506215. S2CID 20328411.
- ^ Vaughan, Martha; Hill, Robert L.; Simoni, Robert D. (2002). "La determinación del fósforo y el descubrimiento de la fosfocreatina y el ATP: el trabajo de Fiske y SubbaRow". Journal of Biological Chemistry . 277 (32): e21. PMID 12161449. Archivado desde el original el 8 de agosto de 2017. Consultado el 24 de octubre de 2017 .
- ^ Maruyama, K. (marzo de 1991). "El descubrimiento del trifosfato de adenosina y el establecimiento de su estructura". J. Hist. Biol . 24 (1): 145–154. doi :10.1007/BF00130477. S2CID 87425890.
- ^ Lipmann, F. (1941). "Generación metabólica y utilización de la energía de enlace de fosfato". Adv. Enzymol . 1 : 99–162. ISSN 0196-7398.
- ^ "Historia: el ATP se descubrió por primera vez en 1929". Premio Nobel de Química 1997. Fundación Nobel . Archivado desde el original el 23 de enero de 2010. Consultado el 26 de mayo de 2010 .
- ^ «El Premio Nobel de Química 1997». Premio Nobel . Archivado desde el original el 24 de octubre de 2017. Consultado el 21 de enero de 2018 .
Enlaces externos
- ATP unido a proteínas en el PDB
- ScienceAid: Energía, ATP y ejercicio
- Entrada de PubChem para trifosfato de adenosina
- Entrada de KEGG para trifosfato de adenosina
