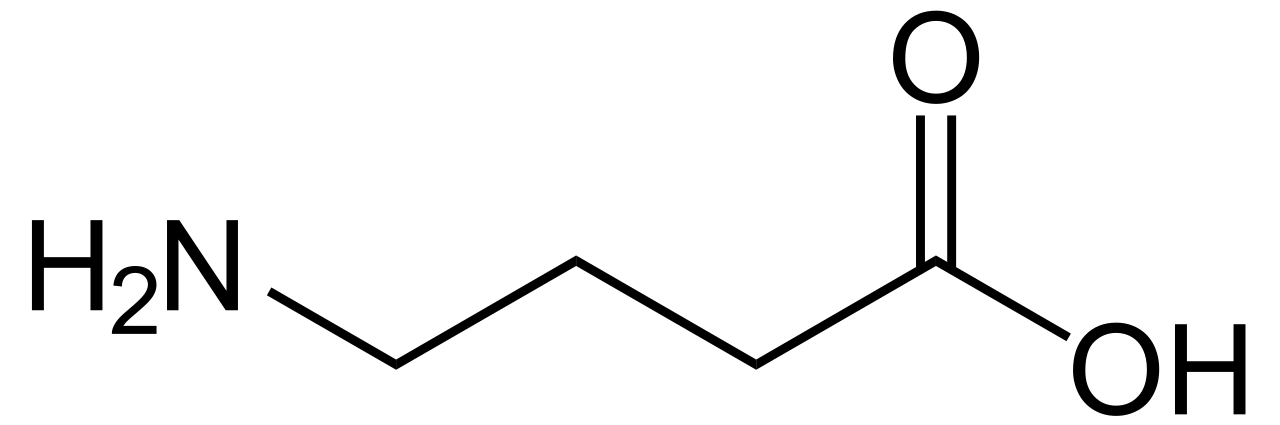
GABA
 | |
 | |
| Nombres | |
|---|---|
| Pronunciación | / ˈ ɡ æ m ə ə ˈ m iː n oʊ b juː ˈ t ɪr ɪ k ˈ æ s ɪ d / , / ˈ ɡ æ b ə / (GABA) |
| Nombre IUPAC preferido Ácido 4-aminobutanoico | |
| Otros nombres Ácido γ-aminobutanoico Ácido 4-aminobutírico Ácido 3-carboxipropilamina Ácido piperídico Ácido piperidínico | |
| Identificadores | |
| |
Modelo 3D ( JSmol ) |
|
| 906818 | |
| EBICh |
|
| Química biológica |
|
| Araña química |
|
| Banco de medicamentos |
|
| Tarjeta informativa de la ECHA | 100.000.235 |
| Número CE |
|
| 49775 | |
| |
| BARRIL |
|
| Malla | Ácido gamma-aminobutírico |
Identificador de centro de PubChem |
|
| Número RTECS |
|
| UNIVERSIDAD |
|
Panel de control CompTox ( EPA ) |
|
| |
| |
| Propiedades | |
| C4H9NO2 | |
| Masa molar | 103,121 g·mol −1 |
| Apariencia | polvo microcristalino blanco |
| Densidad | 1,11 g/ml |
| Punto de fusión | 203,7 °C (398,7 °F; 476,8 K) |
| Punto de ebullición | 247,9 °C (478,2 °F; 521,0 K) |
| 130 gramos/100 ml | |
| registro P | -3,17 |
| Acidez (p K a ) |
|
| Peligros | |
| Seguridad y salud en el trabajo (SST/OHS): | |
Principales peligros | Irritante, nocivo |
| Dosis o concentración letal (LD, LC): | |
LD 50 ( dosis media ) | 12.680 mg/kg (ratón, oral) |
Salvo que se indique lo contrario, los datos se proporcionan para los materiales en su estado estándar (a 25 °C [77 °F], 100 kPa). | |


{kind=link}
{kind=link}
El GABA ( ácido gamma aminobutírico , ácido γ-aminobutírico ) es el principal neurotransmisor inhibidor del sistema nervioso central de los mamíferos en etapa de desarrollo . Su función principal es reducir la excitabilidad neuronal en todo el sistema nervioso .
El GABA se vende como suplemento dietético en muchos países. Tradicionalmente se ha pensado que el GABA exógeno (es decir, tomado como suplemento) no cruza la barrera hematoencefálica , pero los datos obtenidos de investigaciones más recientes (década de 2010) en ratas describen esta noción como poco clara. [2] [3]
La forma carboxilato de GABA es γ-aminobutirato .
Función
Neurotransmisor
Se conocen dos clases generales de receptores GABA : [4]
- GABA A en el que el receptor forma parte de un complejo de canal iónico controlado por ligando [5]
- Receptores metabotrópicos GABA B , que son receptores acoplados a la proteína G que abren o cierran canales iónicos a través de intermediarios ( proteínas G )
{kind=link}

Las neuronas que producen GABA como su salida se denominan neuronas GABAérgicas y tienen principalmente una acción inhibidora en los receptores de los vertebrados adultos. Las células espinosas medianas son un ejemplo típico de células GABAérgicas inhibidoras del sistema nervioso central . Por el contrario, el GABA exhibe acciones tanto excitatorias como inhibidoras en los insectos , mediando la activación muscular en las sinapsis entre los nervios y las células musculares, y también la estimulación de ciertas glándulas . [6] En los mamíferos, algunas neuronas GABAérgicas, como las células de araña , también pueden excitar a sus contrapartes glutamatérgicas. [7] Además de la inhibición fásica de acción rápida, pequeñas cantidades de GABA extracelular pueden inducir una inhibición tónica de escala temporal lenta en las neuronas. [8]
Los receptores GABA A son canales de cloruro activados por ligando: cuando son activados por GABA, permiten el flujo de iones de cloruro a través de la membrana de la célula. [5] El que este flujo de cloruro sea despolarizante (hace que el voltaje a través de la membrana celular sea menos negativo), desviador (no tiene efecto sobre el potencial de membrana de la célula) o inhibidor/hiperpolarizante (hace que la membrana celular sea más negativa) depende de la dirección del flujo de cloruro. Cuando el cloruro neto fluye fuera de la célula, el GABA es despolarizante; cuando el cloruro fluye hacia la célula, el GABA es inhibidor o hiperpolarizante. Cuando el flujo neto de cloruro es cercano a cero, la acción del GABA es desviadora. La inhibición de la derivación no tiene un efecto directo sobre el potencial de membrana de la célula; sin embargo, reduce el efecto de cualquier entrada sináptica coincidente al reducir la resistencia eléctrica de la membrana de la célula. La inhibición de la derivación puede "anular" el efecto excitatorio del GABA despolarizante, lo que resulta en una inhibición general incluso si el potencial de membrana se vuelve menos negativo. Se pensaba que un cambio en el desarrollo de la maquinaria molecular que controla la concentración de cloruro dentro de la célula cambia el papel funcional del GABA entre las etapas neonatal y adulta. A medida que el cerebro se desarrolla hasta la edad adulta, el papel del GABA cambia de excitatorio a inhibidor. [9]
Desarrollo del cerebro
El GABA es un transmisor inhibidor en el cerebro maduro; se pensaba que sus acciones eran principalmente excitatorias en el cerebro en desarrollo. [9] [10] Se informó que el gradiente de cloruro se invierte en neuronas inmaduras, con un potencial de inversión mayor que el potencial de membrana en reposo de la célula; la activación de un receptor GABA-A conduce así a la salida de iones Cl − de la célula (es decir, una corriente despolarizante). Se demostró que el gradiente diferencial de cloruro en neuronas inmaduras se debe principalmente a la mayor concentración de cotransportadores NKCC1 en relación con los cotransportadores KCC2 en células inmaduras. Las interneuronas GABAérgicas maduran más rápido en el hipocampo y la maquinaria GABA aparece antes que la transmisión glutamatérgica. Por lo tanto, se considera que el GABA es el principal neurotransmisor excitatorio en muchas regiones del cerebro antes de la maduración de las sinapsis glutamatérgicas . [11]
En las etapas de desarrollo que preceden a la formación de contactos sinápticos, el GABA es sintetizado por las neuronas y actúa como mediador de señalización tanto autocrino (actúa sobre la misma célula) como paracrino (actúa sobre células cercanas). [12] [13] Las eminencias ganglionares también contribuyen en gran medida a la formación de la población de células corticales GABAérgicas. [14]
El GABA regula la proliferación de células progenitoras neuronales , [15] [16] la migración [17] y la diferenciación [18] [19] la elongación de las neuritas [20] y la formación de sinapsis. [21]
El GABA también regula el crecimiento de células madre neuronales y embrionarias . El GABA puede influir en el desarrollo de células progenitoras neuronales a través de la expresión del factor neurotrófico derivado del cerebro (BDNF). [22] El GABA activa el receptor GABA A , lo que provoca la detención del ciclo celular en la fase S, lo que limita el crecimiento. [23]
Más allá del sistema nervioso
{kind=link}

Además del sistema nervioso, el GABA también se produce en niveles relativamente altos en las células beta productoras de insulina (células β) del páncreas . Las células β secretan GABA junto con la insulina y el GABA se une a los receptores de GABA en las células alfa de los islotes vecinos (células α) y les impide secretar glucagón (que contrarrestaría los efectos de la insulina). [25]
El GABA puede promover la replicación y supervivencia de las células β [26] [27] [28] y también promover la conversión de células α en células β, lo que puede conducir a nuevos tratamientos para la diabetes . [29]
Además de los mecanismos GABAérgicos, también se ha detectado GABA en otros tejidos periféricos, incluidos los intestinos, el estómago, las trompas de Falopio , el útero , los ovarios , los testículos , los riñones , la vejiga urinaria , los pulmones y el hígado , aunque en niveles mucho más bajos que en las neuronas o las células β. [30]
Los experimentos realizados con ratones han demostrado que el hipotiroidismo inducido por intoxicación con flúor se puede detener mediante la administración de GABA. La prueba también demostró que la tiroides se recuperaba de forma natural sin más ayuda después de que el GABA expulsara el flúor. [31]
Las células inmunes expresan receptores para GABA [32] [33] y la administración de GABA puede suprimir las respuestas inmunes inflamatorias y promover respuestas inmunes "reguladoras", de modo que se ha demostrado que la administración de GABA inhibe las enfermedades autoinmunes en varios modelos animales. [26] [32] [34] [35]
En 2018, se ha demostrado que el GABA regula la secreción de un mayor número de citocinas. En el plasma de pacientes con diabetes tipo 1 , los niveles de 26 citocinas están aumentados y, de ellas, 16 son inhibidas por el GABA en los ensayos celulares. [36]
En 2007 se describió un sistema GABAérgico excitatorio en el epitelio de las vías respiratorias . El sistema se activa con la exposición a alérgenos y puede participar en los mecanismos del asma . [37] También se han encontrado sistemas GABAérgicos en los testículos [38] y en el cristalino. [39]
Estructura y conformación
El GABA se encuentra principalmente como un zwitterión (es decir, con el grupo carboxilo desprotonado y el grupo amino protonado). Su conformación depende de su entorno. En la fase gaseosa, una conformación altamente plegada se ve fuertemente favorecida debido a la atracción electrostática entre los dos grupos funcionales. La estabilización es de aproximadamente 50 kcal/mol, según los cálculos de la química cuántica . En el estado sólido, se encuentra una conformación extendida, con una conformación trans en el extremo amino y una conformación gauche en el extremo carboxilo. Esto se debe a las interacciones de empaquetamiento con las moléculas vecinas. En solución, se encuentran cinco conformaciones diferentes, algunas plegadas y otras extendidas, como resultado de los efectos de solvatación . La flexibilidad conformacional del GABA es importante para su función biológica, ya que se ha descubierto que se une a diferentes receptores con diferentes conformaciones. Muchos análogos del GABA con aplicaciones farmacéuticas tienen estructuras más rígidas para controlar mejor la unión. [40] [41]
Historia
En 1883 se sintetizó por primera vez el GABA, que al principio solo se conocía como un producto metabólico de plantas y microbios. [42]
En 1950, los investigadores de la Facultad de Medicina de la Universidad de Washington, Eugene Roberts y Sam Frankel, utilizaron técnicas de cromatografía recientemente desarrolladas para analizar extractos libres de proteínas del cerebro de mamíferos y descubrieron que el GABA se produce a partir del ácido glutámico y se acumula en el sistema nervioso central de los mamíferos . [43] [44]
No hubo mucha más investigación sobre la sustancia hasta siete años después, cuando investigadores canadienses identificaron al GABA como el componente misterioso (denominado Factor I por sus descubridores en 1954) de los extractos de cerebro y médula espinal que inhibía las neuronas del cangrejo de río. [43] [45]
En 1959 se demostró que en una sinapsis inhibidora en las fibras musculares del cangrejo de río, el GABA actúa como estimulación del nervio inhibidor. Tanto la inhibición por estimulación nerviosa como por GABA aplicado son bloqueadas por la picrotoxina . [46]
Biosíntesis
{kind=link}

El GABA se sintetiza principalmente a partir del glutamato a través de la enzima glutamato descarboxilasa (GAD) con fosfato de piridoxal (la forma activa de la vitamina B6 ) como cofactor . Este proceso convierte el glutamato (el principal neurotransmisor excitatorio ) en GABA (el principal neurotransmisor inhibidor). [47] [48]
El GABA también puede sintetizarse a partir de putrescina [49] [50] por la diaminooxidasa y la aldehído deshidrogenasa . [49]
Históricamente se pensaba que el GABA exógeno no penetraba la barrera hematoencefálica [2] , pero investigaciones más actuales [3] describen esta noción como poco clara a la espera de más investigaciones.
Metabolismo
Las enzimas transaminasas GABA catalizan la conversión del ácido 4-aminobutanoico (GABA) y el 2-oxoglutarato (α-cetoglutarato) en semialdehído succínico y glutamato. El semialdehído succínico se oxida luego a ácido succínico por la semialdehído succínico deshidrogenasa y, como tal, entra en el ciclo del ácido cítrico como una fuente utilizable de energía. [51]
Farmacología
Los fármacos que actúan como moduladores alostéricos de los receptores de GABA (conocidos como análogos de GABA o fármacos GABAérgicos ), o que aumentan la cantidad disponible de GABA, suelen tener efectos relajantes, ansiolíticos y anticonvulsivos (con una eficacia equivalente a la de la lamotrigina según estudios realizados en ratones). [52] [53] Se sabe que muchas de las sustancias que se indican a continuación causan amnesia anterógrada y amnesia retrógrada . [54]
En general, el GABA no cruza la barrera hematoencefálica , [2] aunque ciertas áreas del cerebro que no tienen una barrera hematoencefálica efectiva, como el núcleo periventricular , pueden ser alcanzadas por medicamentos como el GABA inyectado sistémicamente. [55] Al menos un estudio sugiere que el GABA administrado por vía oral aumenta la cantidad de hormona de crecimiento humana (HGH). [56] Se ha informado que el GABA inyectado directamente en el cerebro tiene efectos tanto estimulantes como inhibidores en la producción de la hormona de crecimiento, dependiendo de la fisiología del individuo. [55] En consecuencia, considerando los posibles efectos bifásicos del GABA en la producción de la hormona de crecimiento, así como otras preocupaciones de seguridad, no se recomienda su uso durante el embarazo y la lactancia. [57]
El GABA mejora el catabolismo de la serotonina en N -acetilserotonina (el precursor de la melatonina ) en ratas. [58] Por lo tanto, se sospecha que el GABA está involucrado en la síntesis de melatonina y, por lo tanto, podría ejercer efectos reguladores sobre el sueño y las funciones reproductivas. [59]
Química
Aunque en términos químicos el GABA es un aminoácido (ya que tiene tanto un grupo funcional amina primaria como un ácido carboxílico), rara vez se hace referencia a él como tal en la comunidad profesional, científica o médica. Por convención, el término "aminoácido", cuando se utiliza sin un calificativo , se refiere específicamente a un alfa-aminoácido . El GABA no es un alfa-aminoácido, lo que significa que el grupo amino no está unido al carbono alfa. Tampoco se incorpora a las proteínas como lo hacen muchos alfa-aminoácidos. [60]
Fármacos GABAérgicos
Los ligandos del receptor GABA A se muestran en la siguiente tabla [nb 1]
| Actividad en GABA A | Ligando |
|---|---|
| Agonista ortostérico | Muscimol , [61] GABA, [61] gaboxadol ( THIP ), [61] isoguvacina , progabida , ácido piperidina-4-sulfónico (agonista parcial) |
| Moduladores alostéricos positivos | Barbitúricos , [62] benzodiazepinas , [63] esteroides neuroactivos , [64] niacina / niacinamida , [65] no benzodiazepinas (es decir, fármacos z, p. ej., zolpidem ), etomidato , [66] alcohol ( etanol ), [67] [68] [69] metacualona , propofol , estiripentol , [70] y anestésicos [61] (incluidos los anestésicos volátiles ). |
| Antagonista ortostérico (competitivo) | bicuculina , [61] gabazina , [71] tuyona , [72] flumazenil [73] |
| Antagonista no competitivo (por ejemplo, bloqueador de canales) | cicutoxin |
| Moduladores alostéricos negativos | furosemida , enantotoxina , amentoflavona |
Los profármacos GABAérgicos incluyen el hidrato de cloral , que se metaboliza a tricloroetanol , [74] que luego actúa a través del receptor GABA A. [75]
La planta kava contiene compuestos GABAérgicos, entre ellos kavaína , dihidrokavaína , metisticina , dihidrometisticina y yangonina . [76]
 | Esta sección necesita referencias médicas más confiables para su verificación o depende demasiado de fuentes primarias . ( junio de 2015 ) |  |
Otros moduladores GABAérgicos incluyen:
- Ligandos del receptor GABA B. [ cita requerida ]
- Inhibidores de la recaptación de GABA : deramciclano , hiperforina , tiagabina . [ cita requerida ]
- Inhibidores de la transaminasa GABA : gabaculina , fenelzina , valproato , vigabatrina , melisa ( Melissa officinalis ). [78]
- Análogos de GABA : pregabalina , gabapentina , [79] picamilon , progabida [ cita requerida ]
El 4-amino-1-butanol es un precursor bioquímico del GABA y puede convertirse en GABA mediante las acciones de la aldehído reductasa (ALR) y la aldehído deshidrogenasa (ALDH) con γ-aminobutiraldehído (GABAL) como intermediario metabólico . [80]
En las plantas
El GABA también se encuentra en plantas. [81] [82] Es el aminoácido más abundante en el apoplasto de los tomates. [83] La evidencia también sugiere un papel en la señalización celular en plantas. [84] [85]
Véase también
- Ácido 3-aminoisobutírico
- Deficiencia de 4-aminobutirato transaminasa (GABA-transaminasa)
- Análogo de GABA
- Receptor GABA
- Té de GABA
- Potencial despolarizante gigante
- Diplejía espástica , una neuropatología neuromuscular por deficiencia de GABA
- Espasticidad
- Deficiencia de semialdehído succínico deshidrogenasa
- Taurina
Notas
- ^ Se enumeran muchos más ligandos de GABA A en Template:GABA receptor modulators y en GABAA receptor#Ligands
Referencias
- ^ Haynes, William M., ed. (2016). Manual de química y física del CRC (97.ª edición). CRC Press . pp. 5–88. ISBN 978-1498754286.
- ^ abc Kuriyama K, Sze PY (enero de 1971). "Barrera hematoencefálica al ácido H3-γ-aminobutírico en animales normales y tratados con ácido aminooxiacético". Neurofarmacología . 10 (1): 103–108. doi :10.1016/0028-3908(71)90013-X. PMID 5569303.
- ^ ab Boonstra E, de Kleijn R, Colzato LS, Alkemade A, Forstmann BU, Nieuwenhuis S (2015). "Neurotransmisores como complementos alimenticios: los efectos del GABA en el cerebro y el comportamiento". Psicólogo Frontal . 6 : 1520. doi : 10.3389/fpsyg.2015.01520 . PMC 4594160 . PMID 26500584.
- ^ Marescaux, C.; Vergnes, M.; Bernasconi, R. (8 de marzo de 2013). Epilepsia generalizada no convulsiva: atención a los receptores GABA-B. Springer Science & Business Media. ISBN 978-3-7091-9206-1.
- ^ ab Phulera, Swastik; Zhu, Hongtao; Yu, Jie; Claxton, Derek P; Yoder, Nate; Yoshioka, Craig; Gouaux, Eric (25 de julio de 2018). "Estructura crio-EM del receptor GABAA triheteromérico α1β1γ2S sensible a las benzodiazepinas en complejo con GABA". eLife . 7 : e39383. doi : 10.7554/eLife.39383 . ISSN 2050-084X. PMC 6086659 . PMID 30044221.
- ^ Ffrench-Constant RH, Rocheleau TA, Steichen JC, Chalmers AE (junio de 1993). "Una mutación puntual en un receptor GABA de Drosophila confiere resistencia a los insecticidas". Nature . 363 (6428): 449–51. Bibcode :1993Natur.363..449F. doi :10.1038/363449a0. PMID 8389005. S2CID 4334499.
- ^ Szabadics J, Varga C, Molnár G, Oláh S, Barzó P, Tamás G (enero de 2006). "Efecto excitador de células axoaxónicas GABAérgicas en microcircuitos corticales". Ciencia . 311 (5758): 233–235. Código Bib : 2006 Ciencia... 311.. 233S. doi : 10.1126/ciencia.1121325. PMID 16410524. S2CID 40744562.
- ^ Koh, Wuhyun; Kwak, Hankyul; Cheong, Eunji; Lee, C. Justin (26 de julio de 2023). "Regulación del tono GABA y sus funciones cognitivas en el cerebro". Nature Reviews Neuroscience . 24 (9): 523–539. doi :10.1038/s41583-023-00724-7. ISSN 1471-003X. PMID 37495761. S2CID 260201740.
- ^ ab Li K, Xu E (junio de 2008). "El papel y el mecanismo del ácido γ-aminobutírico durante el desarrollo del sistema nervioso central". Neurosci Bull . 24 (3): 195–200. doi :10.1007/s12264-008-0109-3. PMC 5552538 . PMID 18500393.
- ^ Ben-Ari Y, Gaiarsa JL, Tyzio R, Khazipov R (octubre de 2007). "GABA: un transmisor pionero que excita neuronas inmaduras y genera oscilaciones primitivas". Physiol. Rev. 87 ( 4): 1215–1284. doi :10.1152/physrev.00017.2006. PMID 17928584.
- ^ Schousboe, Arne; Sonnewald, Ursula (25 de noviembre de 2016). El ciclo del glutamato/GABA-glutamina: homeostasis de los neurotransmisores de aminoácidos. Springer. ISBN 978-3-319-45096-4.
- ^ Purves D, Fitzpatrick D, Hall WC, Augustine GJ, Lamantia AS, eds. (2007). Neurociencia (4.ª ed.). Sunderland, Mass: Sinauer. págs. 135, recuadro 6D. ISBN 978-0-87893-697-7.
- ^ Jelitai M, Madarasz E (2005). "El papel del GABA en el desarrollo neuronal temprano". GABA en el autismo y trastornos relacionados . Revista internacional de neurobiología. Vol. 71. págs. 27–62. doi :10.1016/S0074-7742(05)71002-3. ISBN 9780123668721. Número de identificación personal 16512345.
- ^ Marín O, Rubenstein JL (noviembre de 2001). "Un largo y notable viaje: migración tangencial en el telencéfalo". Nat. Rev. Neurosci . 2 (11): 780–90. doi :10.1038/35097509. PMID 11715055. S2CID 5604192.
- ^ LoTurco JJ, Owens DF, Heath MJ, Davis MB, Kriegstein AR (diciembre de 1995). "GABA y glutamato despolarizan las células progenitoras corticales e inhiben la síntesis de ADN". Neurona . 15 (6): 1287–1298. doi : 10.1016/0896-6273(95)90008-X . PMID 8845153. S2CID 1366263.
- ^ Haydar TF, Wang F, Schwartz ML, Rakic P (agosto de 2000). "Modulación diferencial de la proliferación en las zonas neocorticales ventricular y subventricular". J. Neurosci . 20 (15): 5764–74. doi :10.1523/JNEUROSCI.20-15-05764.2000. PMC 3823557 . PMID 10908617.
- ^ Behar TN, Schaffner AE, Scott CA, O'Connell C, Barker JL (agosto de 1998). "Respuesta diferencial de las células de la placa cortical y de la zona ventricular al GABA como estímulo migratorio". J. Neurosci . 18 (16): 6378–87. doi :10.1523/JNEUROSCI.18-16-06378.1998. PMC 6793175 . PMID 9698329.
- ^ Ganguly K, Schinder AF, Wong ST, Poo M (mayo de 2001). "El propio GABA promueve el cambio de desarrollo de las respuestas GABAérgicas neuronales de la excitación a la inhibición". Cell . 105 (4): 521–32. doi : 10.1016/S0092-8674(01)00341-5 . PMID 11371348. S2CID 8615968.
- ^ Barbin G, Pollard H, Gaïarsa JL, Ben-Ari Y (abril de 1993). "Participación de los receptores GABAA en el crecimiento de neuronas hipocampales cultivadas". Neurosci. Lett . 152 (1–2): 150–154. doi :10.1016/0304-3940(93)90505-F. PMID 8390627. S2CID 30672030.
- ^ Maric D, Liu QY, Maric I, Chaudry S, Chang YH, Smith SV, Sieghart W, Fritschy JM, Barker JL (abril de 2001). "La expresión de GABA domina la progresión del linaje neuronal en el neocórtex embrionario de rata y facilita el crecimiento de neuritas a través de los canales de Cl−/autorreceptor GABA(A)". J. Neurosci . 21 (7): 2343–60. doi :10.1523/JNEUROSCI.21-07-02343.2001. PMC 6762405 . PMID 11264309.
- ^ Ben-Ari Y (septiembre de 2002). "Acciones excitatorias del GABA durante el desarrollo: la naturaleza de la crianza". Nat. Rev. Neurosci . 3 (9): 728–739. doi :10.1038/nrn920. PMID 12209121. S2CID 8116740.
- ^ Obrietan K, Gao XB, Van Den Pol AN (agosto de 2002). "Las acciones excitatorias del GABA aumentan la expresión del BDNF a través de un mecanismo dependiente de MAPK-CREB: un circuito de retroalimentación positiva en las neuronas en desarrollo". J. Neurophysiol . 88 (2): 1005–15. doi :10.1152/jn.2002.88.2.1005. PMID 12163549.
- ^ Wang DD, Kriegstein AR, Ben-Ari Y (2008). "GABA regula la proliferación de células madre antes de la formación del sistema nervioso". Epilepsy Curr . 8 (5): 137–9. doi :10.1111/j.1535-7511.2008.00270.x. PMC 2566617 . PMID 18852839.
- ^ Popp A, Urbach A, Witte OW, Frahm C (2009). Reh TA (ed.). "Las transcripciones de GAD adultas y embrionarias se regulan espaciotemporalmente durante el desarrollo postnatal en el cerebro de la rata". PLoS ONE . 4 (2): e4371. Bibcode :2009PLoSO...4.4371P. doi : 10.1371/journal.pone.0004371 . PMC 2629816 . PMID 19190758.
- ^ Rorsman P, Berggren PO, Bokvist K, Ericson H, Möhler H, Ostenson CG, Smith PA (1989). "La inhibición de la secreción de glucagón por la glucosa implica la activación de los canales de cloruro del receptor GABA A ". Nature . 341 (6239): 233–6. Bibcode :1989Natur.341..233R. doi :10.1038/341233a0. PMID 2550826. S2CID 699135.
- ^ ab Soltani N, Qiu H, Aleksic M, Glinka Y, Zhao F, Liu R, Li Y, Zhang N, Chakrabarti R, Ng T, Jin T, Zhang H, Lu WY, Feng ZP, Prud'homme GJ, Wang Q (2011). "GABA ejerce efectos protectores y regenerativos sobre las células beta de los islotes y revierte la diabetes". Proc. Nacional. Acad. Ciencia. EE.UU . 108 (28): 11692–7. Código bibliográfico : 2011PNAS..10811692S. doi : 10.1073/pnas.1102715108 . PMC 3136292 . PMID 21709230.
- ^ Tian J, Dang H, Chen Z, Guan A, Jin Y, Atkinson MA, Kaufman DL (2013). "El ácido γ-aminobutírico regula tanto la supervivencia como la replicación de las células β humanas". Diabetes . 62 (11): 3760–5. doi :10.2337/db13-0931. PMC 3806626 . PMID 23995958.
- ^ Purwana I, Zheng J, Li X, Deurloo M, Son DO, Zhang Z, Liang C, Shen E, Tadkase A, Feng ZP, Li Y, Hasilo C, Paraskevas S, Bortell R, Greiner DL, Atkinson M, Prud 'hombre GJ, Wang Q (2014). "GABA promueve la proliferación de células β humanas y modula la homeostasis de la glucosa". Diabetes . 63 (12): 4197–205. doi : 10.2337/db14-0153 . PMID 25008178.
- ^ Ben-Othman N, Vieira A, Courtney M, Record F, Gjernes E, Avolio F, Hadzic B, Druelle N, Napolitano T, Navarro-Sanz S, Silvano S, Al-Hasani K, Pfeifer A, Lacas-Gervais S , Leuckx G, Marroquí L, Thévenet J, Madsen OD, Eizirik DL, Heimberg H, Kerr-Conte J, Pattou F, Mansouri A, Collombat P (2017). "La administración de GABA a largo plazo induce la neogénesis de células similares a beta mediada por células alfa". Celúla . 168 (1–2): 73–85.e11. doi : 10.1016/j.cell.2016.11.002 . PMID 27916274.
- ^ Erdö SL, Wolff JR (febrero de 1990). "Ácido γ-aminobutírico fuera del cerebro de los mamíferos". J. Neurochem . 54 (2): 363–72. doi :10.1111/j.1471-4159.1990.tb01882.x. PMID 2405103. S2CID 86144218.
- ^ Yang H, Xing R, Liu S, Yu H, Li P (2016). "El ácido γ-aminobutírico mejora el hipotiroidismo inducido por flúor en ratones Kunming macho". Ciencias de la vida . 146 : 1–7. doi :10.1016/j.lfs.2015.12.041. PMID 26724496.
- ^ ab Tian J, Chau C, Hales TG, Kaufman DL (1999). "Los receptores GABA A median la inhibición de las respuestas de las células T". J. Neuroimmunol . 96 (1): 21–8. doi :10.1016/s0165-5728(98)00264-1. PMID 10227421. S2CID 3006821.
- ^ Mendu SK, Bhandage A, Jin Z, Birnir B (2012). "Diferentes subtipos de receptores GABA-A se expresan en linfocitos T humanos, de ratón y de rata". PLOS ONE . 7 (8): e42959. Bibcode :2012PLoSO...742959M. doi : 10.1371/journal.pone.0042959 . PMC 3424250 . PMID 22927941.
- ^ Tian J, Lu Y, Zhang H, Chau CH, Dang HN, Kaufman DL (2004). "El ácido gamma-aminobutírico inhibe la autoinmunidad de las células T y el desarrollo de respuestas inflamatorias en un modelo murino de diabetes tipo 1". J. Immunol . 173 (8): 5298–304. doi : 10.4049/jimmunol.173.8.5298 . PMID 15470076.
- ^ Tian J, Yong J, Dang H, Kaufman DL (2011). "El tratamiento oral con GABA regula a la baja las respuestas inflamatorias en un modelo murino de artritis reumatoide". Autoinmunidad . 44 (6): 465–70. doi :10.3109/08916934.2011.571223. PMC 5787624 . PMID 21604972.
- ^ Bhandage AK, Jin Z, Korol SV, Shen Q, Pei Y, Deng Q, Espes D, Carlsson PO, Kamali-Moghaddam M, Birnir B (abril de 2018). "+ Células T y es inmunosupresor en la diabetes tipo 1". eBioMedicine . 30 : 283–294. doi :10.1016/j.ebiom.2018.03.019. PMC 5952354 . PMID 29627388.
- ^ Xiang YY, Wang S, Liu M, Hirota JA, Li J, Ju W, Fan Y, Kelly MM, Ye B, Orser B, O'Byrne PM, Inman MD, Yang X, Lu WY (julio de 2007). "Un sistema GABAérgico en el epitelio de las vías respiratorias es esencial para la sobreproducción de moco en el asma". Nat. Med . 13 (7): 862–7. doi :10.1038/nm1604. PMID 17589520. S2CID 2461757.
- ^ Payne AH, Hardy MH (2007). La célula de Leydig en la salud y la enfermedad . Humana Press. ISBN 978-1-58829-754-9.
- ^ Kwakowsky A, Schwirtlich M, Zhang Q, Eisenstat DD, Erdélyi F, Baranyi M, Katarova ZD, Szabó G (diciembre de 2007). "Las isoformas de GAD exhiben patrones de expresión espaciotemporal distintos en el cristalino en desarrollo del ratón: correlación con Dlx2 y Dlx5". Dev. Dyn . 236 (12): 3532–44. doi : 10.1002/dvdy.21361 . PMID : 17969168. S2CID : 24188696.
- ^ Majumdar D, Guha S (1988). "Conformación, potencial electrostático y patrón farmacofórico de GABA (ácido γ-aminobutírico) y varios inhibidores de GABA". Revista de Estructura Molecular: THEOCHEM . 180 : 125–140. doi :10.1016/0166-1280(88)80084-8.
- ^ Sapse AM (2000). Cálculos de orbitales moleculares para aminoácidos y péptidos . Birkhäuser. ISBN 978-0-8176-3893-1.[ página necesaria ]
- ^ Roth RJ, Cooper JR, Bloom FE (2003). La base bioquímica de la neurofarmacología. Oxford [Oxfordshire]: Oxford University Press. pág. 106. ISBN 978-0-19-514008-8.
- ^ ab Spiering, Martin J. (diciembre de 2018). "El descubrimiento de GABA en el cerebro". Journal of Biological Chemistry . 293 (49): 19159–19160. doi : 10.1074/jbc.cl118.006591 . ISSN 0021-9258. PMC 6295731 . PMID 30530855.
- ^ Roberts, E.; Frankel, S. (noviembre de 1950). "Ácido gamma-aminobutírico en el cerebro: su formación a partir del ácido glutámico". Revista de química biológica . 187 (1): 55–63. ISSN 0021-9258. PMID 14794689.
- ^ Bazemore, AW; Elliott, KAC; Florey, E. (agosto de 1957). "Aislamiento del factor I". Revista de neuroquímica . 1 (4): 334–339. doi :10.1111/j.1471-4159.1957.tb12090.x. ISSN 0022-3042.
- ^ WG Van der Kloot; J. Robbins (1959). "Los efectos del GABA y la picrotoxina en el potencial de unión y la contracción del músculo del cangrejo de río". Experientia . 15 : 36.
- ^ Petroff OA (diciembre de 2002). "GABA y glutamato en el cerebro humano". Neurocientífico . 8 (6): 562–573. doi :10.1177/1073858402238515. PMID 12467378. S2CID 84891972.
- ^ Schousboe A, Waagepetersen HS (2007). "GABA: aspectos homeostáticos y farmacológicos". Gaba y los ganglios basales: de las moléculas a los sistemas . Progreso en la investigación del cerebro. vol. 160, págs. 9-19. doi :10.1016/S0079-6123(06)60002-2. ISBN 978-0-444-52184-2. Número de identificación personal 17499106.
- ^ ab Krantis, Anthony (1 de diciembre de 2000). "GABA en el sistema nervioso entérico de los mamíferos". Fisiología . 15 (6): 284–290. doi :10.1152/physiologyonline.2000.15.6.284. ISSN 1548-9213. PMID 11390928.
- ^ Sequerra, EB; Gardino, P.; Hedin-Pereira, C.; de Mello, FG (11 de mayo de 2007). "Putrescina como fuente importante de GABA en la zona subventricular posnatal de rata". Neurociencia . 146 (2): 489–493. doi : 10.1016/j.neuroscience.2007.01.062. ISSN 0306-4522. PMID 17395389. S2CID 43003476.
- ^ Bown AW, Shelp BJ (septiembre de 1997). "El metabolismo y las funciones del ácido γ-aminobutírico". Plant Physiol . 115 (1): 1–5. doi :10.1104/pp.115.1.1. PMC 158453 . PMID 12223787.
- ^ Foster AC, Kemp JA (febrero de 2006). "Terapéutica del sistema nervioso central basada en glutamato y GABA". Curr Opin Pharmacol . 6 (1): 7–17. doi :10.1016/j.coph.2005.11.005. PMID 16377242.
- ^ Chapouthier G, Venault P (octubre de 2001). "¿Un vínculo farmacológico entre la epilepsia y la ansiedad?". Trends Pharmacol. Sci . 22 (10): 491–3. doi :10.1016/S0165-6147(00)01807-1. PMID 11583788.
- ^ Campagna JA, Miller KW, Forman SA (mayo de 2003). "Mecanismos de acción de los anestésicos inhalados". N. inglés. J. Med . 348 (21): 2110–24. doi :10.1056/NEJMra021261. PMID 12761368.
- ^ ab Müller EE, Locatelli V, Cocchi D (abril de 1999). "Control neuroendocrino de la secreción de la hormona del crecimiento". Physiol. Rev. 79 (2): 511–607. doi :10.1152/physrev.1999.79.2.511. PMID 10221989.
- ^ Powers ME, Yarrow JF, McCoy SC, Borst SE (enero de 2008). "Respuestas de las isoformas de la hormona del crecimiento a la ingestión de GABA en reposo y después del ejercicio". Medicina y ciencia en deportes y ejercicio . 40 (1): 104–10. doi : 10.1249/mss.0b013e318158b518 . PMID 18091016. S2CID 24907247.
- ^ Oketch-Rabah HA, Madden EF, Roe AL, Betz JM (agosto de 2021). "Revisión de seguridad del ácido gamma-aminobutírico (GABA) de la Farmacopea de los Estados Unidos (USP)". Nutrients . 13 (8): 2742. doi : 10.3390/nu13082742 . PMC 8399837 . PMID 34444905.
- ^ Balemans MG, Mans D, Smith I, Van Benthem J (1983). "La influencia del GABA en la síntesis de N-acetilserotonina, melatonina, O-acetil-5-hidroxitriptófalo y O-acetil-5-metoxitriptófalo en la glándula pineal de la rata Wistar macho". Reproducción, nutrición, desarrollo . 23 (1): 151–60. doi : 10.1051/rnd:19830114 . PMID 6844712.
- ^ Sato S, Yinc C, Teramoto A, Sakuma Y, Kato M (2008). "Modulación sexualmente dimórfica de las corrientes del receptor GABA(A) por la melatonina en neuronas de la hormona liberadora de gonadotropina de ratas". The Journal of Physiological Sciences . 58 (5): 317–322. doi : 10.2170/physiolsci.rp006208 . PMID 18834560.
- ^ Hellier, Jennifer L. (16 de diciembre de 2014). El cerebro, el sistema nervioso y sus enfermedades [3 volúmenes]. ABC-CLIO. ISBN 978-1-61069-338-7.
- ^ abcde Chua HC, Chebib M (2017). "Receptores GABA a y diversidad en su estructura y farmacología". Receptores GABAA y diversidad en su estructura y farmacología . Avances en farmacología. Vol. 79. págs. 1–34. doi :10.1016/bs.apha.2017.03.003. ISBN 9780128104132. Número de identificación personal 28528665. Número de identificación personal 41704867.
- ^ Löscher, W.; Rogawski, MA (2012). "Cómo evolucionaron las teorías sobre el mecanismo de acción de los barbitúricos". Epilepsia . 53 : 12–25. doi : 10.1111/epi.12025 . PMID 23205959. S2CID 4675696.
- ^ Olsen RW, Betz H (2006). "GABA y glicina". En Siegel GJ, Albers RW, Brady S, Price DD (eds.). Neuroquímica básica: aspectos moleculares, celulares y médicos (7.ª ed.). Elsevier. págs. 291–302. ISBN 978-0-12-088397-4.
- ^
- Herd MB, Belelli D, Lambert JJ (octubre de 2007). "Modulación neuroesteroides de los receptores GABA(A) sinápticos y extrasinápticos". Farmacología y terapéutica . 116 (1): 20–34. doi :10.1016/j.pharmthera.2007.03.007. PMID 17531325.
- Hosie AM, Wilkins ME, da Silva HM, Smart TG (noviembre de 2006). "Los neuroesteroides endógenos regulan los receptores GABAA a través de dos sitios transmembrana discretos". Nature . 444 (7118): 486–9. Bibcode :2006Natur.444..486H. doi :10.1038/nature05324. PMID 17108970. S2CID 4382394.
- Agís-Balboa RC, Pinna G, Zhubi A, Maloku E, Veldic M, Costa E, Guidotti A (septiembre de 2006). "Caracterización de neuronas cerebrales que expresan enzimas que median la biosíntesis de neuroesteroides". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 103 (39): 14602–7. Bibcode :2006PNAS..10314602A. doi : 10.1073/pnas.0606544103 . PMC 1600006 . PMID 16984997.
- Akk G, Shu HJ, Wang C, Steinbach JH, Zorumski CF, Covey DF, Mennerick S (diciembre de 2005). "Acceso de neurosteroides al receptor GABAA". La Revista de Neurociencia . 25 (50): 11605–13. doi :10.1523/JNEUROSCI.4173-05.2005. PMC 6726021 . PMID 16354918.
- Belelli D, Lambert JJ (julio de 2005). "Neurosteroides: reguladores endógenos del receptor GABA(A)". Nature Reviews. Neuroscience . 6 (7): 565–75. doi :10.1038/nrn1703. PMID 15959466. S2CID 12596378.
- Pinna G, Costa E, Guidotti A (junio de 2006). "La fluoxetina y la norfluoxetina aumentan de forma estereoespecífica y selectiva el contenido de neuroesteroides cerebrales en dosis que son inactivas en la recaptación de 5-HT". Psicofarmacología . 186 (3): 362–72. doi :10.1007/s00213-005-0213-2. PMID 16432684. S2CID 7799814.
- Dubrovsky BO (febrero de 2005). "Esteroides, esteroides neuroactivos y neuroesteroides en psicopatología". Progreso en neuropsicofarmacología y psiquiatría biológica . 29 (2): 169–92. doi :10.1016/j.pnpbp.2004.11.001. PMID 15694225. S2CID 36197603.
- Mellon SH, Griffin LD (2002). "Neurosteroides: bioquímica y significado clínico". Tendencias en endocrinología y metabolismo . 13 (1): 35–43. doi :10.1016/S1043-2760(01)00503-3. PMID 11750861. S2CID 11605131.
- Puia G, Santi MR, Vicini S, Pritchett DB, Purdy RH, Paul SM, Seeburg PH, Costa E (mayo de 1990). "Los neuroesteroides actúan sobre los receptores GABAA humanos recombinantes". Neuron . 4 (5): 759–65. doi :10.1016/0896-6273(90)90202-Q. PMID 2160838. S2CID 12626366.
- Majewska MD, Harrison NL, Schwartz RD, Barker JL, Paul SM (mayo de 1986). "Los metabolitos de las hormonas esteroides son moduladores similares a los barbitúricos del receptor GABA". Science . 232 (4753): 1004–7. Bibcode :1986Sci...232.1004D. doi :10.1126/science.2422758. PMID 2422758.
- Reddy DS, Rogawski MA (2012). "Neurosteroides: reguladores endógenos de la susceptibilidad a las convulsiones y su papel en el tratamiento de la epilepsia". En Noebels JL, Avoli M, Rogawski MA, et al. (eds.). Jasper's Basic Mechanisms of the Epilepsies (4.ª ed.). Bethesda, Maryland: Centro Nacional de Información Biotecnológica. PMID 22787590.
- ^ Toraskar, Mrunmayee; Pratima RP Singh; Shashank Neve (2010). "Estudio de agonistas GABAérgicos" (PDF) . Deccan Journal of Pharmacology . 1 (2): 56–69. Archivado desde el original (PDF) el 2013-10-16 . Consultado el 2019-04-01 .
- ^ Vanlersberghe, C; Camu, F (2008). "Etomidato y otros no barbitúricos". Anestésicos modernos . Manual de farmacología experimental. Vol. 182. págs. 267–82. doi :10.1007/978-3-540-74806-9_13. ISBN 978-3-540-72813-9. Número de identificación personal 18175096.
- ^ Dzitoyeva S, Dimitrijevic N, Manev H (2003). "El receptor 1 del ácido γ-aminobutírico B media las acciones de alteración de la conducta del alcohol en Drosophila: interferencia del ARN adulto y evidencia farmacológica". Proc. Natl. Sci. USA . 100 (9): 5485–5490. Bibcode :2003PNAS..100.5485D. doi : 10.1073/pnas.0830111100 . PMC 154371 . PMID 12692303.
- ^ Mihic SJ, Ye Q, Wick MJ, Koltchine VV, Krasowski MD, Finn SE, Mascia MP, Valenzuela CF, Hanson KK, Greenblatt EP, Harris RA, Harrison NL (1997). "Sitios de acción del alcohol y de los anestésicos volátiles sobre los receptores GABA A y glicina". Nature . 389 (6649): 385–389. Bibcode :1997Natur.389..385M. doi :10.1038/38738. PMID 9311780. S2CID 4393717.
- ^ Fuente no clara. Una de las siguientes:
- Boehm SL, Ponomarev I, Jennings AW, Whiting PJ, Rosahl TW, Garrett EM, Blednov YA, Harris RA (2004). "Ratones mutantes de la subunidad a del receptor del ácido γ-aminobutírico: Nuevas perspectivas sobre las acciones del alcohol". Farmacología bioquímica . 67 (8): 1581–1602. doi :10.1016/j.bcp.2004.07.023. PMID 17175815.
- Boehm SL, Ponomarev I, Blednov YA, Harris RA (2006). "Del gen al comportamiento y viceversa: nuevas perspectivas sobre la selectividad de las subunidades del receptor GABA A en las acciones del alcohol". En Enna SJ (ed.). GABA . Avances en farmacología. Vol. 54. Elsevier. págs. 171–203. doi :10.1016/S1054-3589(06)54008-6. ISBN 978-0-12-032957-1. Número de identificación personal 17175815.
- ^ Fisher JL (enero de 2009). "El anticonvulsivo estiripentol actúa directamente sobre el receptor GABA(A) como modulador alostérico positivo". Neurofarmacología . 56 (1): 190–7. doi :10.1016/j.neuropharm.2008.06.004. PMC 2665930 . PMID 18585399.
- ^ Ueno, S; Bracamontes, J; Zorumski, C; Weiss, DS; Steinbach, JH (1997). "La bicuculina y la gabazina son inhibidores alostéricos de la apertura del canal del receptor GABAA". The Journal of Neuroscience . 17 (2): 625–34. doi :10.1523/jneurosci.17-02-00625.1997. PMC 6573228 . PMID 8987785.
- ^ Olsen RW (abril de 2000). "Receptores de absenta y ácido gamma-aminobutírico". Proc. Natl. Sci. USA . 97 (9): 4417–8. Bibcode :2000PNAS...97.4417O. doi : 10.1073/pnas.97.9.4417 . PMC 34311 . PMID 10781032.
- ^ Whitwam, JG; Amrein, R. (1 de enero de 1995). "Farmacología del flumazenil". Acta Anestesiológica Scandinavica. Suplemento . 108 : 3–14. doi :10.1111/j.1399-6576.1995.tb04374.x. ISSN 0515-2720. PMID 8693922. S2CID 24494744.
- ^ Jira, Reinhard; Kopp, Erwin; McKusick, Blaine C.; Röderer, Gerhard; Bosch, Axel; Fleischmann, Gerald. "Cloroacetaldehídos". Enciclopedia de química industrial de Ullmann . Weinheim: Wiley-VCH. doi :10.1002/14356007.a06_527.pub2. ISBN 978-3527306732.
- ^ Lu, J.; Greco, MA (2006). "Circuitos del sueño y el mecanismo hipnótico de los fármacos GABAA". Journal of Clinical Sleep Medicine . 2 (2): S19–S26. doi : 10.5664/jcsm.26527 . PMID 17557503.
- ^ Singh YN, Singh NN (2002). "Potencial terapéutico de la kava en el tratamiento de los trastornos de ansiedad". CNS Drugs . 16 (11): 731–43. doi :10.2165/00023210-200216110-00002. PMID 12383029. S2CID 34322458.
- ^ Dimitrijevic N, Dzitoyeva S, Satta R, Imbesi M, Yildiz S, Manev H (2005). " Los receptores GABA B de Drosophila están involucrados en los efectos conductuales del ácido gamma-hidroxibutírico (GHB)". Eur. J. Pharmacol . 519 (3): 246–252. doi :10.1016/j.ejphar.2005.07.016. PMID 16129424.
- ^ Awad R, Muhammad A, Durst T, Trudeau VL, Arnason JT (agosto de 2009). "Fraccionamiento guiado por bioensayo de melisa ( Melissa officinalis L.) utilizando una medida in vitro de la actividad de la transaminasa GABA". Phytother Res . 23 (8): 1075–81. doi :10.1002/ptr.2712. PMID 19165747. S2CID 23127112.
- ^ Celikyurt IK, Mutlu O, Ulak G, Akar FY, Erden F (2011). "La gabapentina, un análogo de GABA, mejora el rendimiento cognitivo en ratones". Cartas de Neurociencia . 492 (2): 124–8. doi :10.1016/j.neulet.2011.01.072. PMID 21296127. S2CID 8303292.
- ^ Storer, R. James; Ferrante, Antonio (10 de octubre de 1997). "Ensayo radioquímico de la diamina oxidasa". Protocolos de poliaminas . Métodos en biología molecular. Vol. 79. Nueva Jersey: Humana Press. págs. 91–96. doi :10.1385/0-89603-448-8:91. ISBN. 978-0-89603-448-8. Número de identificación personal 9463822.
- ^ Ramesh SA, Tyerman SD, Xu B, Bose J, Kaur S, Conn V, Domingos P, Ullah S, Wege S, Shabala S, Feijó JA, Ryan PR, Gilliham M, Gillham M (2015). "La señalización de GABA modula el crecimiento de las plantas regulando directamente la actividad de los transportadores de aniones específicos de las plantas". Nat Commun . 6 : 7879. Bibcode :2015NatCo...6.7879R. doi :10.1038/ncomms8879. PMC 4532832 . PMID 26219411.
- ^ Ramesh SA, Tyerman SD, Gilliham M, Xu B (2016). "Señalización del ácido γ-aminobutírico (GABA) en plantas". Cell. Mol. Life Sci . 74 (9): 1577–1603. doi :10.1007/s00018-016-2415-7. hdl : 2440/124330 . PMC: 11107511. PMID : 27838745. S2CID : 19475505.
- ^ Park DH, Mirabella R, Bronstein PA, Preston GM, Haring MA, Lim CK, Collmer A, Schuurink RC (octubre de 2010). "Las mutaciones en los genes de las transaminasas del ácido γ-aminobutírico (GABA) en plantas o Pseudomonas syringae reducen la virulencia bacteriana". Planta J. 64 (2): 318–30. doi : 10.1111/j.1365-313X.2010.04327.x . PMID 21070411.
- ^ Bouché N, Fromm H (marzo de 2004). "GABA en plantas: ¿solo un metabolito?". Trends Plant Sci . 9 (3): 110–5. doi :10.1016/j.tplants.2004.01.006. PMID 15003233.
- ^ Roberts MR (septiembre de 2007). "¿Actúa el GABA como señal en las plantas?: pistas de estudios moleculares". Plant Signal Behav . 2 (5): 408–9. Bibcode :2007PlSiB...2..408R. doi :10.4161/psb.2.5.4335. PMC 2634229 . PMID 19704616.
Enlaces externos
- Smart TG, Stephenson FA (2019). "Medio siglo de ácido γ-aminobutírico". Brain Neurosci Adv . 3 : 2398212819858249. doi :10.1177/2398212819858249. PMC 7058221. PMID 32166183 .
- Parviz M, Vogel K, Gibson KM, Pearl PL (25 de noviembre de 2014). "Trastornos del metabolismo del GABA: deficiencias de SSADH y GABA-transaminasa". Journal of Pediatric Epilepsy . 3 (4): 217–227. doi :10.3233/PEP-14097. PMC 4256671 . PMID 25485164.
Trastornos clínicos que se sabe que afectan al metabolismo hereditario del GABA
- Espectro de espectrometría de masas del ácido gamma-aminobutírico
- Artículo de Scholarpedia sobre GABA
- Lista de neuronas GABA en NeuroLex.org
- Efectos de la administración oral de ácido gamma-aminobutírico (GABA) sobre el estrés y el sueño en humanos: una revisión sistemática
