ATPasa
| Adenosina trifosfatasa | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identificadores | |||||||||
| N.º CE | 3.6.1.3 | ||||||||
| N.º CAS | 9000-83-3 | ||||||||
| Bases de datos | |||||||||
| IntEnz | Vista de IntEnz | ||||||||
| BRENDA | Entrada de BRENDA | ||||||||
| Expasí | Vista de NiceZyme | ||||||||
| BARRIL | Entrada de KEGG | ||||||||
| MetaCiclo | vía metabólica | ||||||||
| PRIAMO | perfil | ||||||||
| Estructuras del PDB | RCSB AP APBE APSUMA | ||||||||
| |||||||||
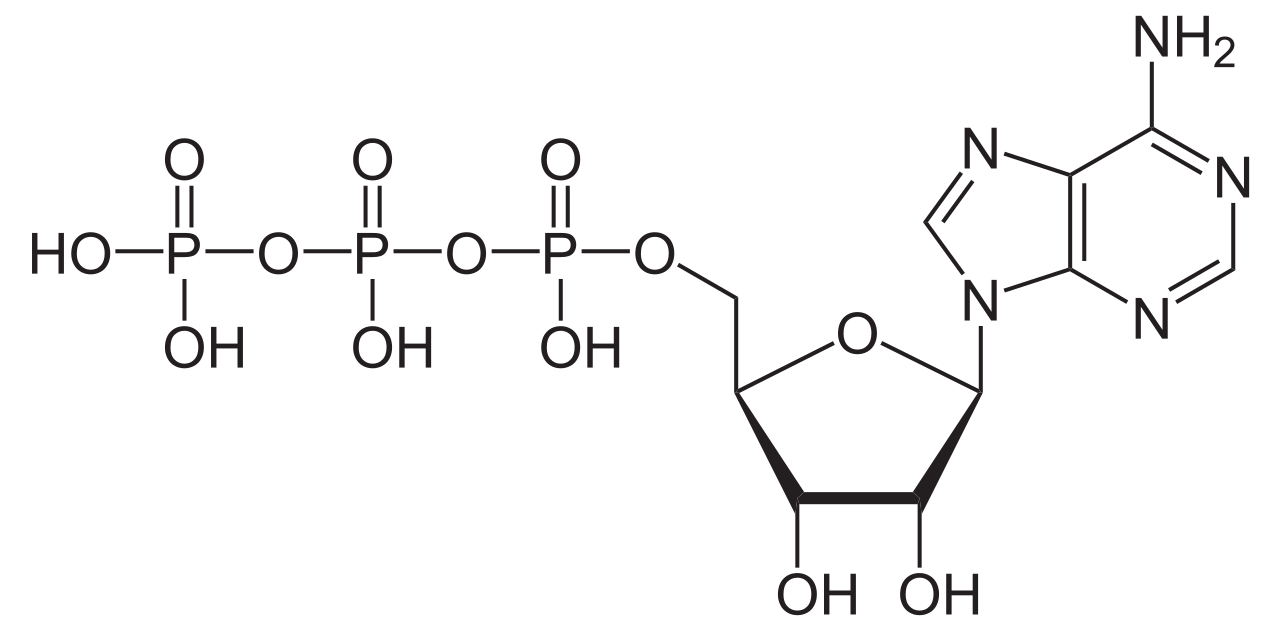

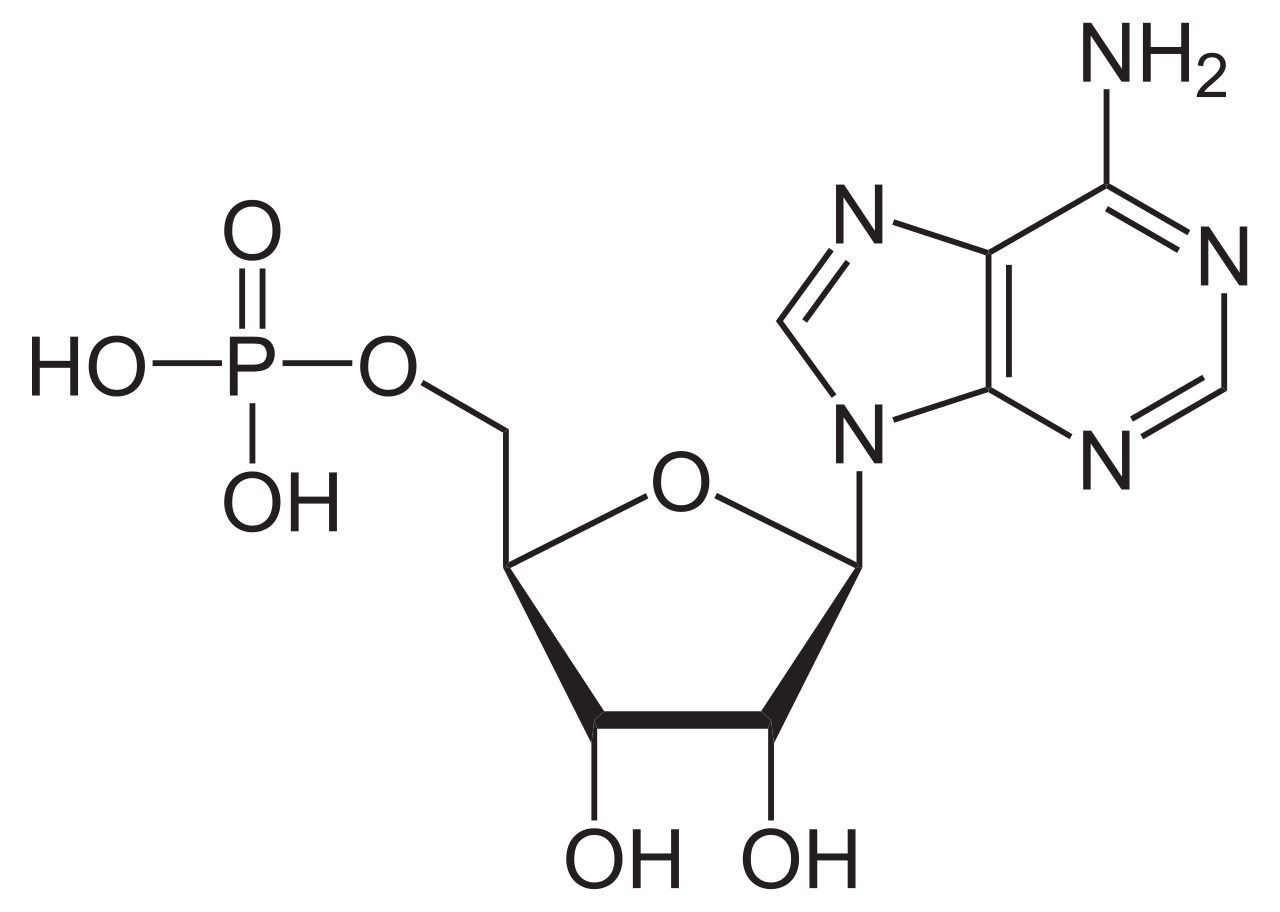
Las ATPasas ( EC 3.6.1.3, A denosina 5'- Trifosfatasa , adenilpirofosfatasa, ATP monofosfatasa, trifosfatasa, antígeno T SV40, ATP hidrolasa, complejo V (transporte de electrones mitocondrial), (Ca 2+ + Mg 2+ )-ATPasa, HCO 3 − -ATPasa , adenosina trifosfatasa) son una clase de enzimas que catalizan la descomposición de ATP en ADP y un ion fosfato libre [1] [2] [ 3] [4] [5] [6] o la reacción inversa. Esta reacción de desfosforilación libera energía , que la enzima (en la mayoría de los casos) aprovecha para impulsar otras reacciones químicas que de otra manera no ocurrirían. Este proceso se usa ampliamente en todas las formas de vida conocidas .
Algunas de estas enzimas son proteínas integrales de membrana (ancladas dentro de las membranas biológicas ) y mueven solutos a través de la membrana, generalmente en contra de su gradiente de concentración. Estas enzimas se denominan ATPasas transmembrana.
Funciones

Las ATPasas transmembrana importan metabolitos necesarios para el metabolismo celular y exportan toxinas, desechos y solutos que pueden obstaculizar los procesos celulares. Un ejemplo importante es la bomba de sodio-potasio (Na + /K + ATPasa) que mantiene el potencial de membrana celular . Otro ejemplo es la ATPasa de hidrógeno y potasio (H + /K + ATPasa o bomba de protones gástrica) que acidifica el contenido del estómago. La ATPasa se conserva genéticamente en los animales; por lo tanto, los cardenólidos , que son esteroides tóxicos producidos por plantas que actúan sobre las ATPasas, forman toxinas animales generales y efectivas que actúan de manera dependiente de la dosis. [7]
Además de los intercambiadores, otras categorías de ATPasa transmembrana incluyen cotransportadores y bombas (sin embargo, algunos intercambiadores también son bombas). Algunos de estos, como la ATPasa Na + /K + , causan un flujo neto de carga, pero otros no. Estos se denominan transportadores electrogénicos y transportadores electroneutrales, respectivamente. [8]
Estructura
Los motivos Walker son un motivo de secuencia de proteínas revelador de la unión de nucleótidos y la hidrólisis. Más allá de esta amplia función, los motivos Walker se pueden encontrar en casi todas las ATPasas naturales, con la notable excepción de las tirosina quinasas . [9] Los motivos Walker forman comúnmente una hélice alfa de hoja Beta -giro- que se autoorganiza como un Nido (motivo estructural de proteína) . Se cree que esto se debe a que las ATPasas modernas evolucionaron a partir de pequeños péptidos de unión a NTP que tuvieron que autoorganizarse. [10]
El diseño de proteínas ha sido capaz de replicar la función de la ATPasa (débilmente) sin utilizar secuencias o estructuras de ATPasa naturales. Es importante destacar que, si bien todas las ATPasas naturales tienen cierta estructura de lámina beta, la "ATPasa alternativa" diseñada carece de estructura de lámina beta, lo que demuestra que esta función esencial para la vida es posible con secuencias y estructuras que no se encuentran en la naturaleza. [11]
Mecanismo
La ATPasa (también llamada F 0 F 1 -ATP sintasa) es un complejo de transferencia de carga que cataliza el ATP para realizar la síntesis de ATP moviendo iones a través de la membrana. [12]
El acoplamiento de la hidrólisis y el transporte de ATP es una reacción química en la que se transporta un número fijo de moléculas de soluto por cada molécula de ATP hidrolizada; para el intercambiador Na + /K + , esto es tres iones Na + fuera de la célula y dos iones K+ dentro por cada molécula de ATP hidrolizada.
Las ATPasas transmembrana utilizan la energía potencial química del ATP realizando trabajo mecánico: transportan solutos en la dirección opuesta a su dirección de movimiento preferida termodinámicamente, es decir, desde el lado de la membrana con baja concentración al lado con alta concentración. Este proceso se conoce como transporte activo .
Por ejemplo, la inhibición de las H + -ATPasas vesiculares daría lugar a un aumento del pH dentro de las vesículas y una caída del pH del citoplasma.
Todas las ATPasas comparten una estructura básica común. Cada ATPasa rotatoria está compuesta por dos componentes principales: F 0 /A 0 /V 0 y F 1 /A 1 /V 1 . Están conectados por 1-3 tallos para mantener la estabilidad, controlar la rotación y evitar que giren en la otra dirección. Un tallo se utiliza para transmitir el par. [13] La cantidad de tallos periféricos depende del tipo de ATPasa: las F-ATPasas tienen uno, las A-ATPasas tienen dos y las V-ATPasas tienen tres. El dominio catalítico F 1 se encuentra en el lado N de la membrana y está involucrado en la síntesis y degradación de ATP y está involucrado en la fosforilación oxidativa . El dominio transmembrana F 0 está involucrado en el movimiento de iones a través de la membrana. [12]
La F 0 F 1 -ATPasa bacteriana consta del dominio soluble F 1 y del dominio transmembrana F 0 , que está compuesto de varias subunidades con estequiometría variable. Hay dos subunidades, γ y ε, que forman el tallo central y están unidas a F 0 . F 0 contiene un oligómero de subunidad c en forma de anillo (anillo c). La subunidad α está cerca de la subunidad b 2 y forma el tallo que conecta las subunidades transmembrana a las subunidades α3β3 y δ. Las F-ATP sintasas son idénticas en apariencia y función, excepto la F 0 F 1 -ATP sintasa mitocondrial, que contiene 7-9 subunidades adicionales. [12]
El potencial electroquímico es lo que hace que el anillo c rote en el sentido de las agujas del reloj para la síntesis de ATP. Esto hace que el tallo central y el dominio catalítico cambien de forma. La rotación del anillo c hace que se formen tres moléculas de ATP, lo que a su vez hace que el H + se mueva del lado P de la membrana al lado N de la misma. La rotación en sentido contrario a las agujas del reloj del anillo c es impulsada por la hidrólisis del ATP y los iones se mueven del lado N al lado P, lo que ayuda a generar potencial electroquímico. [12]
Sintetasas de ATP transmembrana
La ATP sintasa de las mitocondrias y los cloroplastos es una enzima anabólica que aprovecha la energía de un gradiente transmembrana de protones como fuente de energía para agregar un grupo fosfato inorgánico a una molécula de difosfato de adenosina (ADP) para formar una molécula de trifosfato de adenosina (ATP).
Esta enzima funciona cuando un protón se mueve a favor del gradiente de concentración, lo que le da a la enzima un movimiento giratorio. Este movimiento giratorio único une el ADP y el P para crear ATP.
La ATP sintasa también puede funcionar a la inversa, es decir, utilizar la energía liberada por la hidrólisis del ATP para bombear protones contra su gradiente electroquímico.
Clasificación
Existen diferentes tipos de ATPasas, que pueden diferir en función (síntesis y/o hidrólisis de ATP), estructura (las ATPasas F, V y A contienen motores rotatorios) y en el tipo de iones que transportan.
- ATPases rotatorias [14] [15]
- Las F-ATPasas (F1FO-ATPasas) en las mitocondrias , cloroplastos y membranas plasmáticas bacterianas son los principales productores de ATP, utilizando el gradiente de protones generado por la fosforilación oxidativa (mitocondrias) o la fotosíntesis (cloroplastos). [16]
- Las F-ATPasas que carecen de una subunidad delta/OSCP desplazan iones de sodio. Se propone que se las denomine N-ATPasas , ya que parecen formar un grupo distinto que está más separado de las F-ATPasas habituales que las A-ATPasas de las V-ATPasas. [17]
- Las V-ATPasas (V1VO-ATPasas) se encuentran principalmente en vacuolas eucariotas y catalizan la hidrólisis de ATP para transportar solutos y reducir el pH en orgánulos como la bomba de protones del lisosoma.
- Las A-ATPasas (A1AO-ATPasas) se encuentran en las arqueas y algunas bacterias extremófilas. Están organizadas como las V-ATPasas, pero funcionan como las F-ATPasas, principalmente como ATP sintasas.
- Existen muchos homólogos que no son necesariamente rotatorios. Véase ATP sintasa § Evolución .
- Las F-ATPasas (F1FO-ATPasas) en las mitocondrias , cloroplastos y membranas plasmáticas bacterianas son los principales productores de ATP, utilizando el gradiente de protones generado por la fosforilación oxidativa (mitocondrias) o la fotosíntesis (cloroplastos). [16]
- Las P-ATPasas (E1E2-ATPasas) se encuentran en bacterias, hongos y en las membranas plasmáticas y orgánulos eucariotas, y funcionan para transportar una variedad de iones diferentes a través de las membranas.
- Las E-ATPasas son enzimas de la superficie celular que hidrolizan una variedad de NTP, incluido el ATP extracelular. Algunos ejemplos son las ecto-ATPasas, las CD39 y las ecto-ATP/Dasas, todas ellas miembros de una superfamilia "GDA1 CD39". [18]
- Las proteínas AAA son una familia de NTPasas de bucle P con forma de anillo .
P-ATPasa
Las P-ATPasas (conocidas también como E1-E2) se encuentran en bacterias y también en las membranas plasmáticas y organelos eucariotas. Su nombre se debe a la breve unión del fosfato inorgánico a los residuos de aspartato en el momento de la activación. La función de la P-ATPasa es transportar una variedad de compuestos diferentes, como iones y fosfolípidos, a través de una membrana utilizando la hidrólisis de ATP para obtener energía. Existen muchas clases diferentes de P-ATPasas, que transportan un tipo específico de ion. Las P-ATPasas pueden estar compuestas por uno o dos polipéptidos y, por lo general, pueden adoptar dos conformaciones principales, E1 y E2.
Genes humanos
- Transporte de Na + /K + : ATP1A1 , ATP1A2 , ATP1A3 , ATP1A4 , ATP1B1 , ATP1B2, ATP1B3 , ATP1B4
- Transporte de Ca ++ : ATP2A1 , ATP2A2 , ATP2A3 , ATP2B1 , ATP2B2 , ATP2B3 , ATP2B4 , ATP2C1 , ATP2C2
- Transporte de Mg ++ : ATP3
- Intercambio de H + /K + : ATP4A
- Transporte de H + , mitocondrial : ATP5A1 , ATP5B , ATP5C1 , ATP5C2, ATP5D , ATP5E , ATP5F1 , ATP5G1 , ATP5G2 , ATP5G3 , ATP5H , ATP5I , ATP5J , ATP5J2 , ATP5L , ATP5L2, ATP5O , ATP5S , MT-ATP6 , MT-ATP8
- Transporte de H + , lisosómico : ATP6AP1 , ATP6AP2 , ATP6V1A , ATP6V1B1 , ATP6V1B2 , ATP6V1C1 , ATP6V1C2 , ATP6V1D , ATP6V1E1 , ATP6V1E2 , ATP6V1F , ATP6V1G1 , ATP6V1G2 , ATP6V1G3 , ATP6V1H , ATP6V0A1 , ATP6V0A2 , ATP6V0A4 , ATP6V0B , ATP6V0C , ATP6V0D1 , ATP6V0D2 , ATP6V0E
- Transporte de Cu ++ : ATP7A , ATP7B
- Clase I, tipo 8: ATP8A1, ATP8B1 , ATP8B2, ATP8B3 , ATP8B4
- Clase II, tipo 9: ATP9A , ATP9B
- Clase V, tipo 10: ATP10A , ATP10B, ATP10D
- Clase VI, tipo 11: ATP11A, ATP11B , ATP11C
- Transporte de H + /K + , no gástrico: ATP12A
- tipo 13: ATP13A1, ATP13A2 , ATP13A3 , ATP13A4, ATP13A5
Véase también
Referencias
- ^ Geider K, Hoffmann-Berling H (1981). "Proteínas que controlan la estructura helicoidal del ADN". Revista Anual de Bioquímica . 50 : 233–60. doi :10.1146/annurev.bi.50.070181.001313. PMID 6267987.
- ^ Kielley WW (1961). "Myosin adenosine triphosphatase" (Trifosfatasa de adenosina de miosina). En Boyer PD, Lardy H, Myrbäck K (eds.). The Enzymes ( Las enzimas ). Vol. 5 (2.ª ed.). Nueva York: Academic Press. págs. 159-168.
- ^ Martin SS, Senior AE (noviembre de 1980). "Actividades de la adenosina trifosfatasa de membrana en el páncreas de la rata". Biochimica et Biophysica Acta (BBA) - Biomembranas . 602 (2): 401–18. doi :10.1016/0005-2736(80)90320-x. PMID 6252965.
- ^ Njus D, Knoth J, Zallakian M (1981). "Transporte ligado a protones en gránulos cromafines". Temas actuales en bioenergética . 11 : 107–147. doi :10.1016/B978-0-12-152511-8.50010-4.
- ^ Riley MV, Peters MI (junio de 1981). "La localización de la actividad de la ATPasa sensible a los aniones en el endotelio corneal". Biochimica et Biophysica Acta (BBA) - Biomembranas . 644 (2): 251–6. doi :10.1016/0005-2736(81)90382-5. PMID 6114746.
- ^ Tjian R (1981). "Regulación de la transcripción viral y la replicación del ADN por el antígeno T grande de SV40". Temas actuales en microbiología e inmunología . 93 : 5–24. doi :10.1007/978-3-642-68123-3_2. ISBN 978-3-642-68125-7. Número PMID 6269805.
- ^ Dobler S, Dalla S, Wagschal V, Agrawal AA (agosto de 2012). "Evolución convergente a nivel comunitario en la adaptación de los insectos a los cardenólidos tóxicos mediante sustituciones en la Na,K-ATPasa". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 109 (32): 13040–5. doi : 10.1073/pnas.1202111109 . PMC 3420205 . PMID 22826239.
- ^ "3.2: Transporte en membranas". Biology LibreTexts . 21 de enero de 2017 . Consultado el 28 de julio de 2022 .
- ^ Walker JE, Saraste M, Runswick MJ, Gay NJ (1982). "Secuencias distantemente relacionadas en las subunidades alfa y beta de la ATP sintasa, miosina, quinasas y otras enzimas que requieren ATP y un pliegue de unión de nucleótidos común". EMBO J . 1 (8): 945–51. doi :10.1002/j.1460-2075.1982.tb01276.x. PMC 553140 . PMID 6329717.
- ^ Romero Romero ML, Yang F, Lin YR, Toth-Petroczy A, Berezovsky IN, Goncearenco A, et al. (diciembre de 2018). "Proteínas de bucle de fosfato simples pero funcionales". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 115 (51): E11943–E11950. doi : 10.1073/pnas.1812400115 . PMC 6304952 . PMID 30504143.
- ^ Wang M, Hecht MH (agosto de 2020). "Una ATPasa completamente de novo a partir del diseño de proteínas combinatorias". Revista de la Sociedad Química Estadounidense . 142 (36): 15230–15234. doi :10.1021/jacs.0c02954. PMID 32833456.
- ^ abcd Calisto F, Sousa FM, Sena FV, Refojo PN, Pereira MM (febrero de 2021). "Mecanismos de transducción de energía por proteínas de membrana translocadoras de carga". Chemical Reviews . 121 (3): 1804–1844. doi : 10.1021/acs.chemrev.0c00830 . PMID 33398986.
- ^ Hahn A, Parey K, Bublitz M, Mills DJ, Zickermann V, Vonck J, et al. (agosto de 2016). "La estructura de un dímero completo de la ATP sintasa revela la base molecular de la morfología de la membrana mitocondrial interna". Molecular Cell . 63 (3): 445–456. doi : 10.1016/j.molcel.2016.05.037 . PMC 4980432 . PMID 27373333.
- ^ Stewart AG, Laming EM, Sobti M, Stock D (abril de 2014). "ATPases rotatorias: máquinas moleculares dinámicas". Current Opinion in Structural Biology . 25 : 40–8. doi : 10.1016/j.sbi.2013.11.013 . PMID 24878343.
- ^ Kühlbrandt W, Davies KM (enero de 2016). "ATPases rotatorias: una nueva versión de una máquina antigua". Tendencias en ciencias bioquímicas . 41 (1): 106–116. doi :10.1016/j.tibs.2015.10.006. PMID 26671611.
- ^ Watanabe R, Noji H (abril de 2013). "Mecanismo de acoplamiento quimiomecánico de la F(1)-ATPasa: catálisis y generación de par". FEBS Letters . 587 (8): 1030–1035. doi :10.1016/j.febslet.2013.01.063. PMID 23395605.
- ^ Dibrova DV, Galperin MY, Mulkidjanian AY (junio de 2010). "Caracterización de la N-ATPasa, una forma distinta, de transferencia lateral de Na+-translocación de la ATPasa de membrana bacteriana de tipo F". Bioinformática . 26 (12): 1473–1476. doi : 10.1093/bioinformatics/btq234 . PMC 2881411 . PMID 20472544.
- ^ Knowles AF (marzo de 2011). "La superfamilia GDA1_CD39: NTPDases con diversas funciones". Señalización purinérgica . 7 (1): 21–45. doi :10.1007/s11302-010-9214-7. PMC 3083126 . PMID 21484095.
Enlaces externos
- "La ATP sintasa: una espléndida máquina molecular"
- ATPasa en los encabezados de temas médicos (MeSH) de la Biblioteca Nacional de Medicina de EE. UU.
- Estructuras de microscopía electrónica de ATPasas del EM Data Bank (EMDB)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
