Implantación (embriología)
| Implantación | |
|---|---|
 La implantación como una de las primeras etapas del desarrollo embrionario humano | |
| Detalles | |
| Escenario Carnegie | 3 |
| Días | 5–9 |
| Da lugar a | Gástrula |
| Identificadores | |
| Malla | D010064 |
| Terminología anatómica [editar en Wikidata] | |
{kind=link}
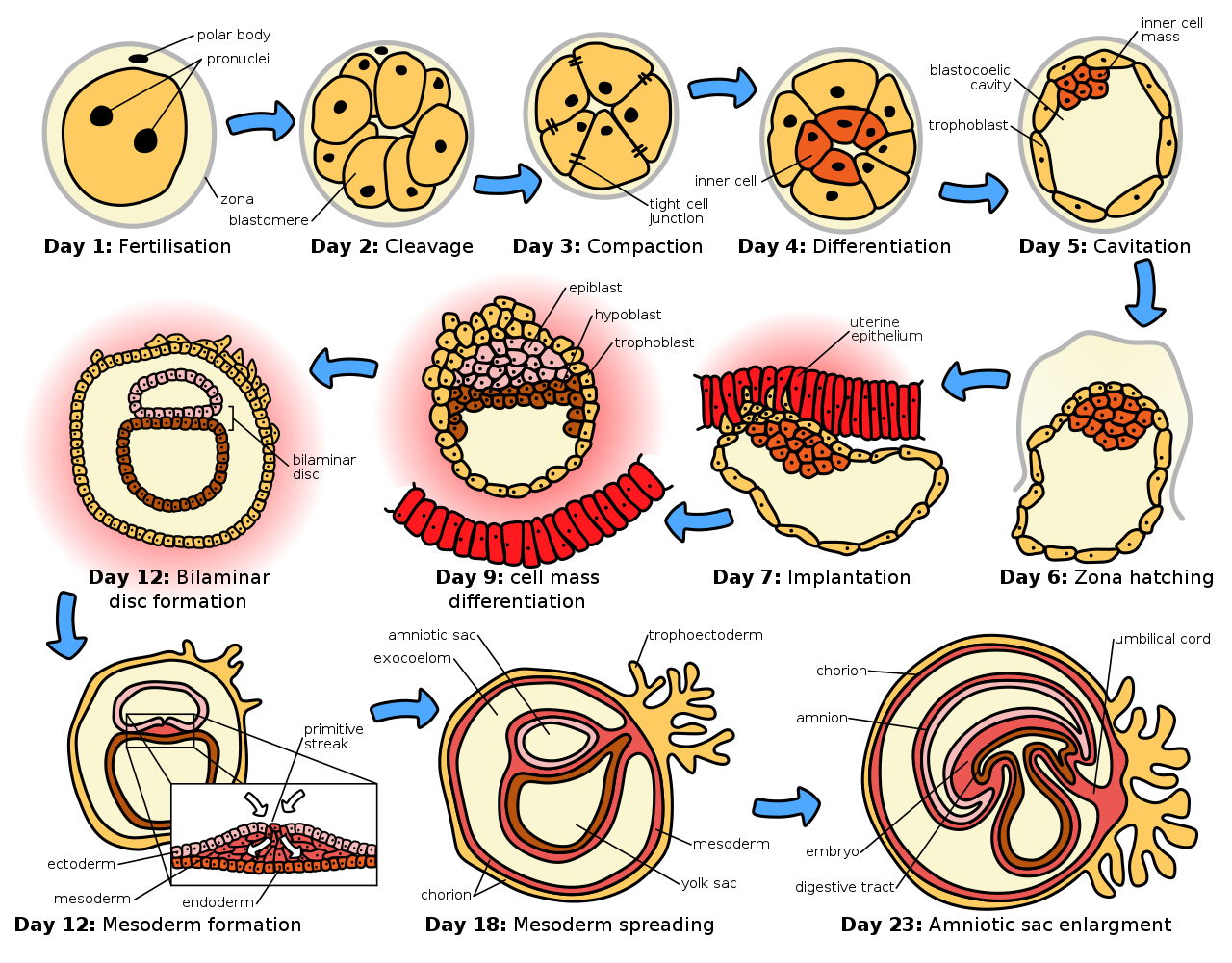
La implantación , también conocida como nidación , [1] es la etapa del desarrollo embrionario de los mamíferos en la que el blastocisto eclosiona, se adhiere, se adhiere e invade el endometrio del útero de la hembra . [2] La implantación es la primera etapa de la gestación y, cuando tiene éxito, se considera que la hembra está embarazada . [3] Un embrión implantado se detecta por la presencia de niveles elevados de gonadotropina coriónica humana (hCG) en una prueba de embarazo . [3] El embrión implantado recibirá oxígeno y nutrientes para crecer.
Para que se produzca la implantación, el útero debe volverse receptivo. La receptividad uterina implica una gran comunicación cruzada entre el embrión y el útero, lo que inicia cambios en el endometrio. Esta etapa genera una sincronía que abre una ventana de implantación que permite la implantación exitosa de un embrión viable. [4] El sistema endocannabinoide desempeña un papel vital en esta sincronía en el útero, influyendo en la receptividad uterina y la implantación del embrión. [5] El embrión expresa receptores cannabinoides en las primeras etapas de su desarrollo que responden a la anandamida (AEA) secretada en el útero. La AEA se produce en niveles más altos antes de la implantación y luego se regula a la baja en el momento de la implantación. Esta señalización es importante en la comunicación cruzada embrión-útero para regular el momento de la implantación embrionaria y la receptividad uterina. Se necesitan concentraciones adecuadas de AEA que no sean ni demasiado altas ni demasiado bajas para una implantación exitosa. [5] [6] [7]
Existe una amplia variación en el tipo de células del trofoblasto y las estructuras de la placenta en las diferentes especies de mamíferos. [8] De las cinco etapas reconocidas de implantación, incluidas dos etapas previas a la implantación que preceden a la placentación , las primeras cuatro son similares en todas las especies. Las cinco etapas son migración y eclosión, precontacto, unión, adhesión e invasión. [8] Las dos etapas previas a la implantación están asociadas con el embrión previo a la implantación . [9] [10]
En los seres humanos, tras la etapa de eclosión que tiene lugar alrededor de cuatro a cinco días después de la fecundación , comienza el proceso de implantación. Al final de la primera semana, el blastocisto se adhiere superficialmente al endometrio uterino. Al final de la segunda semana, la implantación se ha completado. [11]
Etapas de implantación
Hay cinco etapas reconocidas de implantación en mamíferos, incluyendo dos etapas previas a la implantación que preceden a la formación de la placenta . Son: migración y eclosión, precontacto, unión, adhesión e invasión. Las primeras cuatro etapas son similares en todas las especies y el proceso de invasión es variable. [8] [12] Estas tres etapas de aposición , unión e invasión también se denominan alternativamente contacto (aposición), adhesión (unión) y penetración (invasión), [10] [9] y solo pueden tener lugar durante un período de tiempo limitado conocido como la ventana de implantación, cuando el útero está en su punto más receptivo.
Migración y eclosión
{kind=link}

Hay dos etapas de migración involucradas en la implantación, la primera es la migración del cigoto , y la segunda es la migración del trofoblasto . [13] La fertilización del ovocito, tiene lugar en la ampolla de la trompa de Falopio . Los cilios en el revestimiento de la trompa mueven al cigoto en su migración hacia el útero. [13]
Durante esta migración, el cigoto sufre una serie de divisiones celulares que crean una bola de 16 blastómeros compactados llamada mórula . [14] La mórula ingresa al útero después de tres o cuatro días y, mientras lo hace, se forma una cavidad llamada blastocele en la mórula para producir el blastocisto . El blastocisto contiene la masa celular interna que se desarrollará hasta convertirse en el embrión propiamente dicho, y una capa celular externa de trofoblastos que se desarrollará hasta convertirse en las membranas extraembrionarias ( membranas fetales ). [15]
El blastocisto todavía está encerrado en la cubierta del huevo conocida como zona pelúcida , y para poder implantarse en la pared uterina debe deshacerse de esta cubierta. Esta etapa se conoce como eclosión de la zona , y cuando hay suficiente disolución, el blastocisto puede iniciar la etapa de aposición de la implantación. Los factores líticos en la cavidad uterina, así como los factores del propio blastocisto, son esenciales para la ruptura de la cubierta del huevo. Los mecanismos en este último caso están indicados por el hecho de que la zona pelúcida permanece intacta si un óvulo no fertilizado se coloca en el útero en las mismas condiciones. [16]
Entre los reguladores moleculares conocidos que promueven la eclosión se encuentran predominantemente las proteasas que son estimuladas por varios factores de crecimiento . [17] El blastocisto también produce citocinas , tanto proinflamatorias como antiinflamatorias, que tienen papeles cruciales durante la implantación y otras etapas del embarazo. Ambos tipos de citocinas modulan la actividad de las proteasas, incluyendo MMP , activadores del plasminógeno y catepsinas . [17] Se desconoce si las citocinas involucradas en la eclosión son proinflamatorias o antiinflamatorias, o qué proteasas están involucradas. Sin embargo, es bien aceptado que las citocinas proinflamatorias son dominantes durante la implantación. Las citocinas también están presentes en la leche uterina que podría regular el desarrollo y la función del blastocisto, pero no hay evidencia que respalde su participación en la eclosión. El factor inhibidor de la leucemia (LIF) es una citocina proinflamatoria expresada en el endometrio durante la fase lútea del ciclo menstrual , con la expresión más alta observada durante la ventana de implantación. El LIF juega un papel en la adhesión y la invasión. [17]
La eclosión asistida de la zona puede tener lugar en la reproducción asistida, donde la zona pelúcida puede perforarse artificialmente para facilitar la eclosión. [18]
Aposición
Después de la eclosión de la zona, la primera conexión o contacto suelto entre el blastocisto y el endometrio se llama aposición. La aposición generalmente se realiza donde hay una pequeña cripta en el endometrio, y también donde ha habido suficiente ruptura de la zona pelúcida para permitir que el trofoblasto del blastocisto entre en contacto directo con el endometrio subyacente. Finalmente, la masa celular interna (también embrioblasto), dentro de la capa del trofoblasto , se alinea más cerca de la decidua . Si la masa celular interna no está alineada con la decidua en la aposición, tiene la capacidad de rotar libremente dentro del trofoblasto y lograr esta alineación. La aposición es solo una interacción débil del trofectodermo con el epitelio uterino que es inestable al esfuerzo cortante . La aposición también es reversible, lo que permite el reposicionamiento del blastocisto en el útero. [14]
Adhesión
La adhesión es una unión mucho más fuerte con el endometrio que la aposición laxa. [ cita requerida ]
Los trofoblastos se adhieren penetrando el endometrio, con protrusiones de células trofoblásticas. [ cita requerida ]
Esta actividad de adhesión se lleva a cabo mediante microvellosidades que se encuentran en el trofoblasto. El trofoblasto tiene conexiones de fibras de unión, laminina, colágeno tipo IV e integrinas que ayudan en este proceso de adhesión. [19]
La mucina-16 es una mucina transmembrana expresada en la superficie apical de los epitelios uterinos. Esta mucina evita que el blastocisto se implante en un lugar no deseado del epitelio. Por lo tanto, la mucina-16 inhibe la adhesión entre células. Se ha demostrado que su eliminación durante la formación de pinópodos facilita la invasión del trofoblasto in vitro . [20]
La identidad de las moléculas en el trofoblasto y los epitelios endometriales que median la interacción inicial entre los dos sigue sin identificarse. Sin embargo, varios grupos de investigación han propuesto que MUC1 , un miembro de la familia de las mucinas de proteínas glicosiladas, está involucrado. [21] MUC1 es una glicoproteína transmembrana expresada en la superficie apical de las células epiteliales endometriales durante la ventana de implantación en humanos y se ha demostrado que se expresa de manera diferencial entre sujetos fértiles e infértiles durante este tiempo. [21] MUC1 muestra fracciones de carbohidratos en su dominio extracelular que son ligandos de L-selectina , una molécula de adhesión celular en la superficie de las células del trofoblasto. [22] [23] Un modelo in vitro de implantación proporcionó evidencia para apoyar la hipótesis de que L-selectina media la aposición del blastocisto al epitelio uterino al interactuar con sus ligandos. [24]
Invasión
{kind=link}

La invasión es el establecimiento posterior del blastocisto en el endometrio. Las protuberancias de células del trofoblasto que se adhieren al endometrio continúan proliferando y penetrando en el endometrio utilizando gelatinasas A ( MMP-2 ) y B ( MMP-9 ). [25] Los trofoblastos invaden el útero intentando alcanzar el suministro de sangre materna, para establecer la base para el flujo sanguíneo fetal. [26] También secretan factor de preimplantación , un péptido que ayuda a su invasión y formación de placenta. [27] A medida que estos trofoblastos penetran, se fusionan con sus vecinos, diferenciándose terminalmente en un tejido multinucleado , un sincitio conocido como sinciciotrofoblasto . Entre esta capa y el blastocisto se encuentra el citotrofoblasto . [28] [29]
Cuando el sinciciotrofoblasto alcanza la membrana basal debajo de las células deciduales , las desaloja para invadir aún más el estroma uterino. El desalojo se logra degradando las moléculas de adhesión celular (CAM) que unen las células deciduales y la matriz extracelular asociada . La degradación se logra mediante la secreción del factor de necrosis tumoral alfa del sinciciotrofoblasto, que inhibe la expresión de CAM y beta-catenina . La matriz extracelular es degradada por metaloproteinasas como colagenasas , gelatinasas y metaloproteinasas de matriz , y por serina proteasas . [30] Las colagenasas digieren colágeno de tipos I , II , III , VII y X. [30] Las gelatinasas existen en dos formas: una que digiere colágeno de tipo IV y otra que digiere gelatina . [30] La matriz extracelular es degradada por serina endopeptidasas y metaloproteinasas . El sinciciotrofoblasto puede entonces invadir el endometrio llevándose consigo al embrión, donde se incrusta. [30] Finalmente, el sinciciotrofoblasto entra en contacto con la sangre materna y forma vellosidades coriónicas , el comienzo de la placentación . Después de la invasión, la brecha en el epitelio uterino hecha por la entrada del blastocisto se sella con un tapón de fibrina . El tapón de fibrina es una coagulación de un coágulo de sangre y restos celulares. [11]
Trofoblastos extravellosos
Los trofoblastos extravellosos son células de las vellosidades invasoras que migran al miometrio del útero de la madre. Estas células remodelan las arterias espirales para mejorar y asegurar el flujo sanguíneo materno al embrión en crecimiento. También hay evidencia de que este proceso ocurre con las venas uterinas, estabilizándolas para mejorar el drenaje de la sangre fetal y los desechos metabólicos. [31] También se ha documentado que los trofoblastos migran a varios tejidos de la madre. Debido a esto, los trofoblastos han sido implicados en un fenómeno conocido como microquimerismo fetomaterno , donde las células fetales establecen líneas celulares en los tejidos maternos. [32]
Secreciones
Se ha demostrado que los blastocistos preimplantacionales son capaces de secretar factores de crecimiento, hormonas y proteasas similares a la tripsina para participar en el proceso de eclosión. [33]
Durante la invasión, el blastocisto secreta factores para una multitud de propósitos. [33] Secreta varios factores autocrinos , que se dirigen a sí mismos y los estimulan para que invadan aún más el endometrio. La gonadotropina coriónica humana (hCG) es un factor de crecimiento autocrino para el blastocisto, mientras que el factor de crecimiento similar a la insulina 2 estimula su invasividad. [30] La gonadotropina coriónica humana no solo actúa como un inmunosupresor, sino que también le indica a la madre que está embarazada , evitando la luteólisis del cuerpo lúteo y la menstruación al mantener la función del cuerpo lúteo. [30] Las secreciones aflojan las células deciduales entre sí, evitan que la madre rechace al embrión, desencadenan la decidualización final y previenen la menstruación. El factor de preimplantación es secretado por las células del trofoblasto antes de la formación de la placenta. [27]
Inmunosupresor
El embrión es diferente de las células de la madre y, si no secretase agentes inmunosupresores , el sistema inmunitario de la madre lo rechazaría como parásito. Entre estos agentes se encuentran el factor activador de plaquetas , la gonadotropina coriónica humana , el factor de embarazo temprano , la prostaglandina E2 , la interleucina-1 alfa , la interleucina 6 , el interferón alfa, el factor inhibidor de la leucemia y el factor estimulante de colonias . [ cita requerida ]
Otros factores
Otros factores secretados por el blastocisto son: [ cita requerida ]
- Factor liberador de histamina derivado del embrión
- Activador tisular del plasminógeno así como sus inhibidores
- Estradiol
- β1- integrinas
- Factor de crecimiento de fibroblastos
- Factor de crecimiento transformante alfa
- Inhibina
Receptividad del útero
Para permitir la implantación, el útero pasa por cambios para poder recibir el concepto. La receptividad incluye cambios en las células endometriales en la formación de pinópodos que ayudan a absorber el líquido uterino; cambios en el espesor del endometrio y el desarrollo de su irrigación sanguínea, y la formación de la decidua . En conjunto, estos cambios se conocen como transformación de la membrana plasmática y acercan el blastocisto al endometrio y lo inmovilizan. Durante esta etapa, el blastocisto aún puede eliminarse mediante el lavado fuera del útero. [34] [35]
El éxito de la implantación depende de la viabilidad del embrión y de la receptividad del útero. [4] Un factor crítico es la sincronía del desarrollo entre el embrión y el útero. [36] La sincronía proporciona un breve período de receptividad conocido como ventana de implantación, e implica una gran interacción entre el blastocisto y el endometrio en esta etapa. [37] [38] [39]
El sistema endocannabinoide desempeña un papel vital en esta sincronía en el útero, influyendo en la receptividad uterina y la implantación del embrión. [5] El embrión expresa receptores cannabinoides en las primeras etapas de su desarrollo que responden a la anandamida (AEA) secretada en el útero. Esta señalización es importante en la comunicación entre el embrión y el útero para regular el momento de la implantación embrionaria y la receptividad uterina. Se necesitan concentraciones adecuadas de AEA que no sean ni demasiado altas ni demasiado bajas para una implantación exitosa. [5] [40] La IL-6 y la FAAH son cruciales para la receptividad uterina y, junto con la AEA, se observa que existe un vínculo con el grosor endometrial adecuado que sustenta el embarazo. [5]
Durante la adhesión, la comunicación cruzada se transmite por interacciones receptor - ligando , tanto de integrina-matriz como de proteoglicanos. Los receptores de proteoglicanos se encuentran en la superficie de la decidua, y sus contrapartes, los proteoglicanos, se encuentran alrededor de las células del trofoblasto del blastocisto. Este sistema ligando-receptor también está presente justo en la ventana de implantación. [30] El blastocisto envía señales al endometrio para que se adapte más a su presencia, por ejemplo mediante cambios en el citoesqueleto de las células deciduales . Esto, a su vez, desaloja las células deciduales de su conexión con la lámina basal subyacente , lo que permite al blastocisto realizar la invasión posterior. [30]
Ventana de implantación
La ventana de implantación es un período de tiempo limitado para la unión exitosa del blastocisto. [41] En los seres humanos, la receptividad uterina es óptima en los días 20 a 24 de la fase secretora del ciclo menstrual , cuando los niveles de hormona luteinizante están en su punto máximo. [9] [42] La comunicación cruzada entre el embrión y el endometrio tiene lugar durante este tiempo. [9] Las células epiteliales endoteliales que recubren el útero son las primeras células que detectan señales del blastocisto y se transducen en vías de señalización posteriores. [33] En los seres humanos, la ventana de implantación solo está disponible durante 24 a 36 horas. [43]
Se ha señalado que el microbioma endometrial tiene un papel importante en el éxito de la implantación, ya que controla la función de las células endometriales y la función del sistema inmunitario local que impide el crecimiento de patógenos. Esto está asociado con la secreción de sustancias protectoras. [44] [45]
Pinópodos
Los pinópodos se forman al comienzo de la ventana de implantación y se encuentran en muchas especies. [46] [41] Son protuberancias similares a hongos de la membrana celular apical de las células epiteliales uterinas. [41] Los pinópodos se forman por la hinchazón de estas células epiteliales y la fusión de una serie de microvellosidades para alcanzar un tamaño máximo. [46] Aparecen entre el día 19 y el día 21 de la edad gestacional y se forman completamente el día 20. [41] Esto corresponde a una edad de fertilización de aproximadamente cinco a siete días, que se corresponde bien con el momento de la implantación. Los pinópodos solo persisten durante un máximo de dos días y se consideran marcadores ultraestructurales de receptividad. [46]
Su desarrollo es potenciado por la progesterona e inhibido por los estrógenos . Durante la ventana de implantación, la adhesión de célula a célula es inhibida por MUC1 , una glicoproteína de la superficie celular que pertenece al glicocáliz . Los pinópodos son más altos que las microvellosidades y sobresalen a través del glicocáliz, lo que permite el contacto directo con el trofoblasto adherido. El atributo más importante de los pinópodos es esta eliminación de glicoproteínas de las superficies celulares de las células epiteliales uterinas. [9] También se ha demostrado que MUC16 desaparece de las superficies celulares con el desarrollo de los pinópodos. Algunos estudios han informado que los pinópodos atrapan los cilios, lo que impide el movimiento del embrión, y durante la implantación permite el contacto cercano y la adherencia del embrión. [41]
Los pinópodos introducen el líquido uterino y sus macromoléculas en las células mediante el proceso de endocitosis . Esto disminuye el volumen del útero, acercando las paredes al blastocisto que flota en él. Por lo tanto, el período de actividad de los pinópodos podría limitar la ventana de implantación. [30] Los pinópodos continúan absorbiendo líquido, eliminando la mayor parte durante las primeras etapas de la implantación. [47]
Predecidualización
El endometrio aumenta de grosor, se vasculariza y sus glándulas se vuelven tortuosas y se multiplican en sus secreciones. Estos cambios alcanzan su máximo alrededor de siete días después de la ovulación . [ cita requerida ]
Además, la superficie del endometrio produce una especie de células redondeadas, que cubren toda la zona hacia la cavidad uterina. Esto ocurre alrededor de 9 a 10 días después de la ovulación. [30] Estas células se llaman células deciduales , lo que enfatiza que toda la capa de ellas se desprende en cada menstruación si no se produce un embarazo, al igual que las hojas de los árboles de hoja caduca . Las glándulas uterinas, por otro lado, disminuyen su actividad y se degeneran alrededor de 8 a 9 días [30] después de la ovulación en ausencia de embarazo.
Las células deciduales se originan a partir de las células del estroma que siempre están presentes en el endometrio, y forman una nueva capa, la decidua . El resto del endometrio, además, expresa diferencias entre el lado luminal y el basal. Las células luminales forman el estrato compacto del endometrio, en contraste con el estrato esponjoso basalolateral , que consiste en las células del estroma más bien esponjosas. [30]
Decidualización
{kind=link}

La decidualización se expande si se produce un embarazo, lo que permite un mayor desarrollo de las glándulas uterinas, la zona compacta y el epitelio de células deciduales que la recubren. Las células deciduales se llenan de lípidos y glucógeno y adoptan la forma poliédrica característica de las células deciduales. Los factores del blastocisto también desencadenan la formación final de las células deciduales en su forma adecuada. Por el contrario, algunas células deciduales en la proximidad del blastocisto se degeneran, lo que le proporciona nutrientes. [30] Un indicio de la influencia embrionaria es que la decidualización se produce en mayor grado en los ciclos de concepción que en los ciclos de no concepción. [30] Además, se observan cambios similares cuando se dan estímulos que imitan la invasión natural del embrión. [30]
El embrión libera serina proteasas que provocan la despolarización de la membrana de la célula epitelial y activan el canal de sodio epitelial . Esto desencadena una afluencia de iones de calcio (Ca 2+ ) y la fosforilación de CREB. La fosforilación de CREB regula positivamente la expresión de COX2 , lo que conduce a la liberación de prostaglandina E2 (PGE2) de las células epiteliales. La PGE2 actúa sobre las células del estroma activando las vías relacionadas con el AMPc en la célula del estroma que conducen a la decidualización. [48]
Partes de la decidua
La decidua se puede organizar en secciones separadas, aunque tengan la misma composición.
- Decidua basal: es la parte de la decidua que se ubica basalolateral al embrión después de la implantación.
- Decidua capsularis: la decidua capsularis crece sobre el embrión en el lado luminal, encerrándolo en el endometrio y rodea al embrión junto con la decidua basal.
- Decidua parietal: todas las demás deciduas en la superficie uterina pertenecen a la decidua parietal.
Decidua durante el embarazo
Después de la implantación, la decidua permanece, al menos durante el primer trimestre. [30] Sin embargo, su momento más destacado es durante las primeras etapas del embarazo, durante la implantación. Su función como tejido circundante es reemplazada por la placenta definitiva . Sin embargo, algunos elementos de la decidualización permanecen durante todo el embarazo. [30]
Durante el embarazo, las capas compacta y esponjosa todavía se pueden observar debajo de la decidua. Las glándulas de la capa esponjosa continúan secretando durante el primer trimestre, cuando se degeneran. Sin embargo, antes de esa desaparición, algunas glándulas secretan de manera desigual. Este fenómeno de hipersecreción se denomina fenómeno de Arias-Stella [30] , en honor al patólogo Javier Arias-Stella .
Glándulas uterinas
{kind=link}

| Proteínas, glicoproteínas y péptidos secretada por las glándulas uterinas [30] |
| Matriz asociada: |
| Fibronectina |
| Laminina |
| Entactina |
| Colágeno tipo IV |
| Sulfato de heparán |
| Proteoglicano |
| Integrinas |
| – |
| Otros: |
| Mucinas |
| Prolactina |
| IGFBP -1 |
| Glicodelina |
| Proteína endometrial 15 |
| Albúmina |
| Lipoproteína beta |
| Relajación |
| Factor de crecimiento de fibroblastos 1 |
| Factor de crecimiento de fibroblastos 2 |
| Papalisina-1 |
| Proteína de respuesta al estrés 27 (SRP-27) |
| CA-125 |
| Beta-endorfina |
| Leu- encefalina |
| Diaminooxidasa |
| Activador tisular del plasminógeno |
| Renina |
| Anhidrasa carbónica dependiente de progesterona |
| Lactoferrina |
No sólo se transforma el revestimiento del útero, sino que también cambia la secreción de sus glándulas . Este cambio es inducido por el aumento de los niveles de progesterona del cuerpo lúteo . El objetivo de las secreciones es el embrioblasto, y tiene varias funciones sobre él.
Alimento
El embrión pasa aproximadamente 72 horas en la cavidad uterina antes de implantarse. Durante ese tiempo, no puede recibir nutrición directamente de la sangre de la madre y debe depender de nutrientes secretados en la cavidad uterina, por ejemplo, hierro y vitaminas liposolubles. [30]
Crecimiento e implantación
Además de la nutrición, el endometrio secreta varias proteínas dependientes de esteroides , importantes para el crecimiento y la implantación. También se secretan colesterol y esteroides. [30] La implantación se facilita aún más mediante la síntesis de sustancias de la matriz , moléculas de adhesión y receptores de superficie para las sustancias de la matriz.
Importancia clínica
Fallo de implantación
La reproducción en humanos no es muy eficiente. Sólo alrededor del 30% de las concepciones naturales resultan en embarazos exitosos. De los embarazos fallidos, alrededor del 85% se deben a un fallo de implantación. [49] Se considera que el fallo de implantación se debe a una receptividad uterina inadecuada en dos tercios de los casos, y a problemas con el embrión en el otro tercio. [50] La mayoría de los procedimientos de FIV fracasan debido a un fallo de implantación, lo que representa casi la mitad de todos los embarazos fallidos. [49]
La receptividad uterina inadecuada puede ser causada por una señalización hormonal y de citocinas anormal , así como por alteraciones epigenéticas . [51] El fallo recurrente de implantación es una causa de infertilidad femenina . Por lo tanto, las tasas de embarazo se pueden mejorar optimizando la receptividad endometrial para la implantación. [51] La evaluación de los marcadores de implantación puede ayudar a predecir el resultado del embarazo y detectar una deficiencia oculta de implantación. [51] Como parte del programa de órgano en un chip , se ha desarrollado un endometrio en un chip para modelar el funcionamiento del endometrio que podría identificar más claramente las causas del fallo de implantación. [52] También se han desarrollado organoides para modelar el endometrio y su papel en la implantación. [53]
En mujeres con más de tres fallos de implantación en reproducción asistida , una revisión de varios estudios controlados aleatorizados pequeños estimó que el uso de heparina de bajo peso molecular adyuvante mejora la tasa de nacidos vivos en aproximadamente un 80%. [54] El apoyo de la fase lútea puede incluir el uso de progesterona y gonadotropina coriónica humana (hCG) para mejorar las posibilidades de una implantación exitosa. [55]
Deficiencia de zinc
El zinc es crucial en la etapa previa a la concepción (y en el embarazo exitoso), y su deficiencia puede provocar un desarrollo deficiente del blastocisto. Una vez que se fertiliza un óvulo, el zinc se libera en una chispa de zinc que promueve cambios que incluyen el endurecimiento de la zona pelúcida, lo que previene la poliespermia . [56]
Sangrado de implantación
El sangrado y el manchado son comunes durante la fase lútea del ciclo menstrual y las primeras etapas del embarazo, pero no están relacionados con la implantación. El sangrado de implantación ocurre entre 7 y 14 días después de la fertilización, [57] y es una pequeña cantidad de sangrado vaginal leve o manchado que puede ocurrir al comienzo del embarazo debido a que el blastocisto penetra el revestimiento del útero durante la implantación. [58] [59] [60] Para el día 13, el sitio de penetración en el endometrio generalmente se ha cerrado con un tapón de fibrina , pero el aumento del flujo sanguíneo hacia los espacios del sinciciotrofoblasto a veces puede causar sangrado en ese sitio. [58] El sangrado de implantación puede estar acompañado de síntomas como calambres, náuseas, dolor en los senos y dolores de cabeza. [61] El sangrado de implantación se puede distinguir del sangrado menstrual por el color, la coagulación, la fuerza y la duración del flujo. [62] [63]
Véase también
- Cannabinoides
- Diapausa embrionaria
- Células de Hofbauer
- Capacidad homeostática
- Superpoblación humana
- Membrana de Reichert
Referencias
- ^ "Definición de implantación". www.merriam-webster.com . Consultado el 26 de abril de 2023 .
- ^ Niringiyumukiza JD, Cai H, Xiang W (mayo de 2018). "Participación de la prostaglandina E2 en la fertilidad femenina de los mamíferos: ovulación, fertilización, desarrollo embrionario e implantación temprana". Biología reproductiva y endocrinología . 16 (1): 43. doi : 10.1186/s12958-018-0359-5 . PMC 5928575. PMID 29716588 .
- ^ ab Wilcox AJ, Harmon Q, Doody K, Wolf DP, Adashi EY (abril de 2020). "Pérdida preimplantacional de óvulos humanos fertilizados: estimación de lo no observable". Reproducción humana . 35 (4): 743–750. doi :10.1093/humrep/deaa048. PMC 8287936 . PMID 32296829.
- ^ ab Yang Y, Zhu QY, Liu JL (noviembre de 2021). "Descifrando la receptividad uterina del ratón para la implantación de embriones con resolución de célula única". Proliferación celular . 54 (11): e13128. doi :10.1111/cpr.13128. PMC 8560620 . PMID 34558134.
- ^ abcde Ezechukwu HC, Diya CA, Shrestha N, Hryciw DH (septiembre de 2020). "El papel de los endocannabinoides en el embarazo temprano: avances recientes y efectos del consumo de cannabis". Revista estadounidense de fisiología. Endocrinología y metabolismo . 319 (3): E557–E561. doi :10.1152/ajpendo.00210.2020. PMID 32744098. S2CID 220943305.
- ^ Maccarrone M, Valensise H, Bari M, Lazzarin N, Romanini C, Finazzi-Agrò A (2000). "Relación entre la disminución de las concentraciones de anandamida hidrolasa en linfocitos humanos y el aborto espontáneo". Lancet . 355 (9212): 1326–9. doi :10.1016/S0140-6736(00)02115-2. PMID 10776746. S2CID 39733100.
- ^ Dennedy MC, Friel AM, Houlihan DD, Broderick VM, Smith T, Morrison JJ (enero de 2004). "Cannabinoides y el útero humano durante el embarazo" (PDF) . American Journal of Obstetrics and Gynecology . 190 (1): 2–9, discusión 3A. doi :10.1016/j.ajog.2003.07.013. PMID 14749627.
- ^ abc Imakawa K, Bai R, Fujiwara H, Kusama K (enero de 2016). "Implantación y placentación del concepto: moléculas relacionadas con la transición epitelial-mesenquimal, localización de los linfocitos, retrovirus endógenos y exosomas". Medicina y biología reproductiva . 15 (1): 1–11. doi :10.1007/s12522-015-0215-7. PMC 5715838 . PMID 29259417.
- ^ abcde Kim SM, Kim JS (diciembre de 2017). "Una revisión de los mecanismos de implantación". Desarrollo y reproducción . 21 (4): 351–359. doi :10.12717/DR.2017.21.4.351. PMC 5769129 . PMID 29359200.
- ^ ab McGowen MR, Erez O, Romero R, Wildman DE (2014). "La evolución de la implantación del embrión". Revista Internacional de Biología del Desarrollo . 58 (2–4): 155–161. doi :10.1387/ijdb.140020dw. PMC 6053685 . PMID 25023681.
- ^ ab Moore KL (2020). El ser humano en desarrollo: embriología con orientación clínica (undécima edición). Edimburgo. págs. 35–42. ISBN 978-0-323-61154-1.
{{cite book}}: Mantenimiento de CS1: falta la ubicación del editor ( enlace ) - ^ Cakmak H, Taylor HS (marzo de 2011). "Fallo de implantación: mecanismos moleculares y tratamiento clínico". Actualización sobre reproducción humana . 17 (2): 242–253. doi :10.1093/humupd/dmq037. PMC 3039220. PMID 20729534 .
- ^ ab Ochoa-Bernal MA, Fazleabas AT (marzo de 2020). "Eventos fisiológicos de la implantación y decidualización de embriones en primates humanos y no humanos". Revista internacional de ciencias moleculares . 21 (6): 1973. doi : 10.3390/ijms21061973 . PMC 7139778 . PMID 32183093.
- ^ ab Gauster M, Moser G, Wernitznig S, Kupper N, Huppertz B (junio de 2022). "Desarrollo temprano del trofoblasto humano: de la morfología a la función". Ciencias de la vida celular y molecular . 79 (6): 345. doi :10.1007/s00018-022-04377-0. PMC 9167809 . PMID 35661923.
- ^ Sadler TW (2010). Embriología médica de Langman (11.ª ed.). Filadelfia: Lippincott William & Wilkins. pág. 45. ISBN 978-0-7817-9069-7.
- ^ Boro 2004, pág. 1172.
- ^ abc Seshagiri PB, Vani V, Madhulika P (marzo de 2016). "Citocinas y eclosión de blastocistos". American Journal of Reproductive Immunology . 75 (3): 208–17. doi : 10.1111/aji.12464 . PMID 26706391. S2CID 11540123.
- ^ FIV-infertility.com
- ^ Singh H, Aplin JD (julio de 2009). "Moléculas de adhesión en el epitelio endometrial: integridad tisular e implantación del embrión". Journal of Anatomy . 215 (1): 3–13. doi :10.1111/j.1469-7580.2008.01034.x. PMC 2714633 . PMID 19453302.
- ^ Gipson IK, Blalock T, Tisdale A, Spurr-Michaud S, Allcorn S, Stavreus-Evers A, et al. (enero de 2008). "MUC16 se pierde de la superficie del uterodomo (pinópodo) del endometrio humano receptivo: evidencia in vitro de que MUC16 es una barrera para la adherencia del trofoblasto". Biology of Reproduction . 78 (1): 134–142. doi : 10.1095/biolreprod.106.058347 . PMID 17942799. S2CID 44999196.
- ^ ab Margarit L, Taylor A, Roberts MH, Hopkins L, Davies C, Brenton AG, et al. (diciembre de 2010). "MUC1 como discriminador entre el endometrio de pacientes fértiles e infértiles con SOP y endometriosis". The Journal of Clinical Endocrinology and Metabolism . 95 (12): 5320–5329. doi : 10.1210/jc.2010-0603 . PMID 20826587.
- ^ Carson DD, Julian J, Lessey BA, Prakobphol A, Fisher SJ (septiembre de 2006). "MUC1 es un andamiaje para los ligandos de selectina en el útero humano". Frontiers in Bioscience . 11 (1): 2903–2908. doi : 10.2741/2018 . PMID 16720361.
- ^ Francis LW, Yao SN, Powell LC, Griffiths S, Berquand A, Piasecki T, et al. (febrero de 2021). "La MUC1 altamente glicosilada media la unión de alta afinidad de la L-selectina en la superficie endometrial humana". Journal of Nanobiotechnology . 19 (1): 50. doi : 10.1186/s12951-021-00793-9 . PMC 7890821 . PMID 33596915.
- ^ Genbacev OD, Prakobphol A, Foulk RA, Krtolica AR, Ilic D, Singer MS, et al. (enero de 2003). "Adhesión mediada por L-selectina del trofoblasto en la interfase materno-fetal". Science . 299 (5605): 405–408. Bibcode :2003Sci...299..405G. doi :10.1126/science.1079546. PMID 12532021. S2CID 15462671.
- ^ Zhu JY, Pang ZJ, Yu YH (2012). "Regulación de la invasión del trofoblasto: el papel de las metaloproteinasas de matriz". Reseñas en Obstetricia y Ginecología . 5 (3–4): e137–e143. PMC 3594863 . PMID 23483768.
- ^ Carter AM, Enders AC, Pijnenborg R (marzo de 2015). "El papel del trofoblasto invasivo en la implantación y placentación de primates". Philosophical Transactions of the Royal Society of London. Serie B, Ciencias Biológicas . 370 (1663): 20140070. doi :10.1098/rstb.2014.0070. PMC 4305171. PMID 25602074 .
- ^ ab Zare F, Seifati SM, Dehghan-Manshadi M, Fesahat F (mayo de 2020). "Factor de Preimplantación (PIF): un péptido con diversas funciones". Reproducción asistida JBRA . 24 (2): 214–218. doi :10.5935/1518-0557.20190082. PMC 7169918 . PMID 32202400.
- ^ Gardiner C, Tannetta DS, Simms CA, Harrison P, Redman CW, Sargent IL (2011). "Las microvesículas de sincitiotrofoblasto liberadas de placentas de preeclampsia exhiben una mayor actividad del factor tisular". PLOS ONE . 6 (10): e26313. Bibcode :2011PLoSO...626313G. doi : 10.1371/journal.pone.0026313 . PMC 3194796 . PMID 22022598.
- ^ Cindrova-Davies T, Sferruzzi-Perri AN (1 de noviembre de 2022). "Desarrollo y función de la placenta humana". Seminarios en biología celular y del desarrollo . 131 : 66–77. doi : 10.1016/j.semcdb.2022.03.039 . PMID 35393235.
- ^ abcdefghijklmnopqrstu Boron W , Boulpaep E (2004). Fisiología médica: un enfoque celular y molecular . Oxford : Elsevier . ISBN 1-4160-2328-3.OCLC 61527528 .[ página necesaria ]
- ^ Moser G, Weiss G, Sundl M, Gauster M, Siwetz M, Lang-Olip I, et al. (marzo de 2017). "Los trofoblastos extravellosos invaden más que las arterias uterinas: evidencia de la invasión de las venas uterinas". Histoquímica y biología celular . 147 (3): 353–366. doi :10.1007/s00418-016-1509-5. PMC 5344955 . PMID 27774579.
- ^ Dawe GS, Tan XW, Xiao ZC (enero de 2007). "Migración celular del bebé a la madre". Adhesión celular y migración . 1 (1): 19–27. doi :10.4161/cam.4082. PMC 2633676. PMID 19262088 .
- ^ abc Hennes A, Devroe J, De Clercq K, Ciprietti M, Held K, Luyten K, et al. (abril de 2023). "Las secreciones de proteasa por el blastocisto invasor inducen oscilaciones de calcio en las células epiteliales endometriales a través del receptor 2 activado por proteasa". Reprod Biol Endocrinol . 21 (1): 37. doi : 10.1186/s12958-023-01085-7 . PMC 10105462 . PMID 37060079.
- ^ Murphy CR (agosto de 2004). "Receptividad uterina y transformación de la membrana plasmática". Cell Research . 14 (4): 259–267. doi : 10.1038/sj.cr.7290227 . PMID 15353123. S2CID 24023502.
- ^ Murphy CR, Shaw TJ (diciembre de 1994). "Transformación de la membrana plasmática: una respuesta común de las células epiteliales uterinas durante el período periimplantacional". Cell Biology International . 18 (12): 1115–1128. doi :10.1006/cbir.1994.1038. PMID 7703952. S2CID 22769575.
- ^ Teh WT, McBain J, Rogers P (noviembre de 2016). "¿Cuál es la contribución de la asincronía embrionaria-endometrial al fracaso de la implantación?". Journal of Assisted Reproduction and Genetics . 33 (11): 1419–1430. doi :10.1007/s10815-016-0773-6. PMC 5125144 . PMID 27480540.
- ^ Sternberg AK, Buck VU, Classen-Linke I, Leube RE (agosto de 2021). "Cómo las fuerzas mecánicas cambian el endometrio humano durante el ciclo menstrual en preparación para la implantación del embrión". Cells . 10 (8): 2008. doi : 10.3390/cells10082008 . PMC 8391722 . PMID 34440776.
- ^ Wu HM, Chen LH, Hsu LT, Lai CH (noviembre de 2022). "Tolerancia inmunitaria a la implantación embrionaria y al embarazo: el papel de las vesículas extracelulares derivadas de células estromales deciduales humanas y de embriones". Int J Mol Sci . 23 (21): 13382. doi : 10.3390/ijms232113382 . PMC 9658721 . PMID 36362169.
- ^ Liang J, Wang S, Wang Z (noviembre de 2017). "El papel de los microARN en la implantación del embrión". Reprod Biol Endocrinol . 15 (1): 90. doi : 10.1186/s12958-017-0309-7 . PMC 5699189 . PMID 29162091.
- ^ Maccarrone M, Valensise H, Bari M, Lazzarin N, Romanini C, Finazzi-Agrò A (2000). "Relación entre la disminución de las concentraciones de anandamida hidrolasa en linfocitos humanos y el aborto espontáneo". Lancet . 355 (9212): 1326–9. doi :10.1016/S0140-6736(00)02115-2. PMID 10776746. S2CID 39733100.
- ^ abcde Quinn KE, Matson BC, Wetendorf M, Caron KM (febrero de 2020). "Pinopodes: avances recientes, perspectivas actuales y direcciones futuras". Endocrinología molecular y celular . 501 : 110644. doi :10.1016/j.mce.2019.110644. PMC 6962535. PMID 31738970 .
- ^ Xiao Y, Sun X, Yang X, Zhang J, Xue Q, Cai B, et al. (junio de 2010). "El factor inhibidor de la leucemia está desregulado en el endometrio y el líquido de lavado uterino de pacientes con adenomiosis durante la ventana de implantación". Fertilidad y esterilidad . 94 (1): 85–89. doi : 10.1016/j.fertnstert.2009.03.012 . PMID 19361790.
- ^ Ng SW, Norwitz GA, Pavlicev M, Tilburgs T, Simón C, Norwitz ER (junio de 2020). "Decidualización endometrial: el principal impulsor de la salud del embarazo". Int J Mol Sci . 21 (11): 4092. doi : 10.3390/ijms21114092 . PMC 7312091 . PMID 32521725.
- ^ Crha I, Ventruba P, Žáková J, Ješeta M, Pilka R, Lousová E, et al. (2019). "Microbioma uterino y receptividad endometrial". Ceská Ginecología . 84 (1): 49–54. PMID 31213058.
- ^ Moreno I, Codoñer FM, Vilella F, Valbuena D, Martinez-Blanch JF, Jimenez-Almazán J, et al. (Diciembre 2016). "Evidencia de que la microbiota endometrial tiene un efecto sobre el éxito o fracaso de la implantación". American Journal of Obstetrics and Gynecology . 215 (6): 684–703. doi : 10.1016/j.ajog.2016.09.075 . PMID 27717732.
- ^ abc Zhao Y, He D, Zeng H, Luo J, Yang S, Chen J, et al. (septiembre de 2021). "Expresión y significado de miR-30d-5p y SOCS1 en pacientes con fallo recurrente de implantación durante la ventana de implantación". Reprod Biol Endocrinol . 19 (1): 138. doi : 10.1186/s12958-021-00820-2 . PMC 8425163 . PMID 34496883.
- ^ Sharma A, Kumar P (enero de 2012). "Entender la ventana de implantación, un fenómeno crucial". J Hum Reprod Sci . 5 (1): 2–6. doi : 10.4103/0974-1208.97777 . PMC 3409914 . PMID 22870007. (Retractado, ver doi :10.4103/0974-1208.165155, PMID 26538866. )
- ^ Ruan YC, Guo JH, Liu X, Zhang R, Tsang LL, Dong JD, et al. (julio de 2012). "La activación del canal de Na+ epitelial desencadena la liberación y producción de prostaglandina E₂ necesaria para la implantación del embrión". Nature Medicine . 18 (7): 1112–1117. doi :10.1038/nm.2771. PMID 22729284. S2CID 5116022.
- ^ ab Wang Y, Jiang X, Jia L, Wu X, Wu H, Wang Y, et al. (2022). "Una caracterización unicelular de embriones humanos postimplantación cultivados in vitro delinea la morfogénesis en la sincitialización primaria". Frontiers in Cell and Developmental Biology . 10 : 835445. doi : 10.3389/fcell.2022.835445 . PMC 9240912 . PMID 35784461.
- ^ Melford SE, Taylor AH, Konje JC (2013). "De ratones y mujeres: factores que influyen en la implantación exitosa, incluidos los endocannabinoides". Human Reproduction Update . 20 (3): 415–428. doi : 10.1093/humupd/dmt060 . PMID 24306146.
- ^ abc Cakmak H, Taylor HS (2010). "Fallo de implantación: mecanismos moleculares y tratamiento clínico". Actualización sobre reproducción humana . 17 (2): 242–253. doi :10.1093/humupd/dmq037. PMC 3039220. PMID 20729534.
- ^ Ahn J, Yoon MJ, Hong SH, Cha H, Lee D, Koo HS, et al. (septiembre de 2021). "Endometrio vascularizado en un chip diseñado mediante microingeniería tridimensional". Reproducción humana . 36 (10): 2720–2731. doi :10.1093/humrep/deab186. PMC 8450871 . PMID 34363466.
- ^ Rawlings TM, Makwana K, Tryfonos M, Lucas ES (julio de 2021). "Organoides para modelar el endometrio: implantación y más allá". Reprod Fertil . 2 (3): R85–R101. doi :10.1530/RAF-21-0023. PMC 8801025 . PMID 35118399.
- ^ Potdar N, Gelbaya TA, Konje JC, Nardo LG (2013). "Heparina de bajo peso molecular complementaria para mejorar la tasa de nacidos vivos después de un fallo recurrente de implantación: una revisión sistemática y un metanálisis". Human Reproduction Update . 19 (6): 674–684. doi : 10.1093/humupd/dmt032 . PMID 23912476.
- ^ Farquhar C, Marjoribanks J (agosto de 2018). "Tecnología de reproducción asistida: una descripción general de las revisiones Cochrane". Base de datos Cochrane de revisiones sistemáticas . 2018 (8): CD010537. doi : 10.1002/14651858.CD010537.pub5. PMC 6953328. PMID 30117155.
- ^ Garner TB, Hester JM, Carothers A, Diaz FJ (mayo de 2021). "El papel del zinc en la reproducción femenina". Biología de la reproducción . 104 (5): 976–994. doi :10.1093/biolre/ioab023. PMC 8599883 . PMID 33598687.
- ^ "Sangrado de implantación vs. período menstrual: cómo diferenciarlo". ReUnite Rx . 2020-12-09 . Consultado el 2021-07-29 .
- ^ ab Sadler TW (2010). Embriología médica de Langman (11.ª ed.). Filadelfia: Lippincott William & Wilkins. págs. 50-51. ISBN 978-0-7817-9069-7.
- ^ Sheiner E (2011). Sangrado durante el embarazo: una guía completa. Nueva York: Springer. pág. 9. ISBN 978-1-4419-9810-1.
- ^ Harville EW , Wilcox AJ, Baird DD, Weinberg CR (septiembre de 2003). "Sangrado vaginal en el embarazo muy temprano". Reproducción humana . 18 (9): 1944–1947. doi : 10.1093/humrep/deg379 . PMID 12923154.
- ^ Heger, L. Demosthenes E (ed.). "¿Cómo se sienten los calambres de implantación? Cómo diferenciar entre calambres de implantación y calambres menstruales". Insider . Consultado el 29 de julio de 2021 .
- ^ Aggarwal K (2020). "Sangrado vaginal en el embarazo temprano". En Sharma A (ed.). Sala de partos, urgencias . Singapur: Springer. págs. 155-161. doi :10.1007/978-981-10-4953-8_18. ISBN 978-981-10-4953-8.S2CID212812977 .
- ^ "¿Cómo puedo saber si se trata de un sangrado de implantación?". MedicineNet . Consultado el 29 de julio de 2021 .
Libros
- Boron W , Boulpaep E (2004). Fisiología médica: un enfoque celular y molecular . Oxford : Elsevier . ISBN 1-4160-2328-3.OCLC 61527528 .
Lectura adicional
- "Implantación del blastocisto..."
- "Implantación"
Enlaces externos
- Embrión humano virtual que detalla las etapas de desarrollo [ enlace muerto permanente ]
