Neutrófilo
| Neutrófilo | |
|---|---|
.png/1280px-Blausen_0676_Neutrophil_(crop).png) Representación 3D de un neutrófilo | |
 Neutrófilos con núcleos segmentados rodeados de eritrocitos y plaquetas . En el citoplasma se observan gránulos intracelulares ( tinción de Giemsa ). | |
| Detalles | |
| Sistema | Sistema inmunitario |
| Función | Fagocitosis |
| Identificadores | |
| Malla | D009504 |
| EL | H2.00.04.1.02012 |
| FMA | 62860 |
| Términos anatómicos de microanatomía [editar en Wikidata] | |
Los neutrófilos son un tipo de glóbulo blanco fagocítico y forman parte de la inmunidad innata . Más específicamente, forman el tipo más abundante de granulocitos y constituyen entre el 40% y el 70% de todos los glóbulos blancos en los seres humanos. [1] Sus funciones varían en diferentes animales. [2] También se les conoce como neutrocitos, heterófilos o leucocitos polimorfonucleares.
Se forman a partir de células madre en la médula ósea y se diferencian en subpoblaciones de neutrófilos asesinos y neutrófilos enjaulados. Tienen una vida corta (entre 5 y 135 horas, ver § Duración de vida) y son muy móviles, ya que pueden entrar en partes del tejido donde otras células/moléculas no pueden. Los neutrófilos pueden subdividirse en neutrófilos segmentados y neutrófilos en banda (o bandas ). Forman parte de la familia de células polimorfonucleares (PMN) junto con los basófilos y los eosinófilos . [3] [4] [5]
El nombre neutrófilo deriva de las características de tinción en preparaciones histológicas o citológicas con hematoxilina y eosina ( H&E ) . Mientras que los glóbulos blancos basófilos se tiñen de azul oscuro y los glóbulos blancos eosinófilos se tiñen de rojo brillante, los neutrófilos se tiñen de un rosa neutro. Normalmente, los neutrófilos contienen un núcleo dividido en 2 a 5 lóbulos. [6]
Los neutrófilos son un tipo de fagocito y normalmente se encuentran en el torrente sanguíneo . Durante la fase inicial ( aguda ) de la inflamación , particularmente como resultado de una infección bacteriana , exposición ambiental [7] y algunos cánceres, [8] [9] los neutrófilos son una de las primeras células inflamatorias que responden y migran hacia el sitio de la inflamación. Migran a través de los vasos sanguíneos y luego a través del espacio intersticial, siguiendo señales químicas como la interleucina-8 (IL-8), C5a , fMLP , leucotrieno B4 y peróxido de hidrógeno (H 2 O 2 ) [10] en un proceso llamado quimiotaxis . Son las células predominantes en el pus , lo que explica su apariencia blanquecina/amarillenta. [11]
Los neutrófilos son reclutados al sitio de la lesión en cuestión de minutos después del traumatismo y son el sello distintivo de la inflamación aguda. [12] No solo desempeñan un papel central en la lucha contra la infección, sino que también contribuyen al dolor en el período agudo al liberar citocinas proinflamatorias y otros mediadores que sensibilizan a los nociceptores, lo que lleva a una mayor percepción del dolor. [13] Sin embargo, debido a que algunos patógenos son indigeribles, es posible que no puedan resolver ciertas infecciones sin la ayuda de otros tipos de células inmunes.
Estructura


Cuando se adhieren a una superficie, los granulocitos neutrófilos tienen un diámetro promedio de 12 a 15 micrómetros (μm) en frotis de sangre periférica . En suspensión, los neutrófilos humanos tienen un diámetro promedio de 8,85 μm. [14]
Junto con los eosinófilos y los basófilos , forman la clase de células polimorfonucleares , llamadas así por la forma multilobulada del núcleo (en comparación con los linfocitos y monocitos , los otros tipos de glóbulos blancos). El núcleo tiene una apariencia lobulada característica, los lóbulos separados están conectados por cromatina . El nucléolo desaparece a medida que el neutrófilo madura, algo que sucede solo en unos pocos otros tipos de células nucleadas. [15] : 168 Hasta el 17% de los núcleos de neutrófilos humanos femeninos tienen un apéndice en forma de baqueta que contiene el cromosoma X inactivado . [16] En el citoplasma, el aparato de Golgi es pequeño, las mitocondrias y los ribosomas son escasos y el retículo endoplasmático rugoso está ausente. [15] : 170 El citoplasma también contiene alrededor de 200 gránulos, de los cuales un tercio son azurófilos . [15] : 170
Los neutrófilos mostrarán una segmentación creciente (muchos segmentos del núcleo) a medida que maduran. Un neutrófilo normal debe tener de 3 a 5 segmentos. La hipersegmentación no es normal, pero se produce en algunos trastornos, sobre todo en la deficiencia de vitamina B12 . Esto se observa en una revisión manual del frotis de sangre y es positivo cuando la mayoría o todos los neutrófilos tienen 5 o más segmentos.

Los neutrófilos son los glóbulos blancos más abundantes en el cuerpo humano (se producen aproximadamente 10 11 al día); representan aproximadamente el 50-70% de todos los glóbulos blancos (leucocitos). El rango normal establecido para los recuentos sanguíneos humanos varía entre laboratorios, pero un recuento de neutrófilos de 2,5-7,5 × 10 9 /L es un rango normal estándar. Las personas de ascendencia africana y del Medio Oriente pueden tener recuentos más bajos, que siguen siendo normales. [17] Un informe puede dividir los neutrófilos en neutrófilos segmentados y bandas .
Cuando circulan en el torrente sanguíneo y se inactivan, los neutrófilos son esféricos. Una vez activados, cambian de forma y se vuelven más amorfos o similares a amebas y pueden extender seudópodos mientras buscan antígenos . [18]
La capacidad de los neutrófilos para fagocitar bacterias se reduce cuando se ingieren azúcares simples como glucosa, fructosa, sacarosa, miel y jugo de naranja, mientras que la ingestión de almidones no tuvo ningún efecto. El ayuno, por otro lado, fortaleció la capacidad fagocítica de los neutrófilos para fagocitar bacterias. Se concluyó que la función, y no el número, de los fagocitos en la fagocitación de bacterias se alteraba por la ingestión de azúcares. [19] En 2007, investigadores del Instituto Whitehead de Investigación Biomédica descubrieron que dada una selección de azúcares en las superficies microbianas, los neutrófilos reaccionaban a algunos tipos de azúcares de manera preferencial. Los neutrófilos fagocitaban y mataban preferentemente a los objetivos beta-1,6-glucanos en comparación con los objetivos beta-1,3-glucanos. [20] [21]
Desarrollo
Esperanza de vida
Se ha informado, mediante diferentes enfoques, que la vida media de los neutrófilos humanos inactivados en la circulación es de entre 5 y 135 horas. [22] [23]
Tras la activación, se marginan (se colocan adyacentes al endotelio de los vasos sanguíneos) y experimentan una captura dependiente de selectina seguida de una adhesión dependiente de integrina en la mayoría de los casos, después de lo cual migran a los tejidos, donde sobreviven durante 1 a 2 días. [24] También se ha demostrado que los neutrófilos se liberan a la sangre desde una reserva esplénica después de un infarto de miocardio . [25]
La relación de distribución de neutrófilos en la médula ósea, la sangre y el tejido conectivo es de 28:1:25.
Los neutrófilos son mucho más numerosos que los fagocitos monocitos / macrófagos de vida más larga . Es probable que un patógeno (microorganismo o virus causante de enfermedades) se encuentre primero con un neutrófilo. Algunos expertos plantean la hipótesis de que la corta vida de los neutrófilos es una adaptación evolutiva . La corta vida de los neutrófilos minimiza la propagación de aquellos patógenos que parasitan fagocitos (por ejemplo, Leishmania [26] ) porque cuanto más tiempo pasen estos parásitos fuera de una célula huésped , más probabilidades habrá de que sean destruidos por algún componente de las defensas del cuerpo. Además, debido a que los productos antimicrobianos de los neutrófilos también pueden dañar los tejidos del huésped , su corta vida limita el daño al huésped durante la inflamación . [24]
Los neutrófilos serán eliminados después de la fagocitosis de los patógenos por los macrófagos. La PECAM-1 y la fosfatidilserina en la superficie celular están involucradas en este proceso.
Función
Quimiotaxis
Los neutrófilos experimentan un proceso llamado quimiotaxis a través del movimiento ameboide , que les permite migrar hacia sitios de infección o inflamación. Los receptores de la superficie celular permiten a los neutrófilos detectar gradientes químicos de moléculas como la interleucina-8 (IL-8), el interferón gamma (IFN-γ), C3a, C5a y el leucotrieno B4 , que estas células utilizan para dirigir el camino de su migración.
Los neutrófilos tienen una variedad de receptores específicos, incluidos los del complemento , citocinas como las interleucinas y el IFN-γ, quimiocinas , lectinas y otras proteínas. También expresan receptores para detectar y adherirse al endotelio y receptores Fc para la opsonina . [27]
En los leucocitos que responden a un quimioatrayente , la polaridad celular está regulada por las actividades de las pequeñas guanosina trifosfatasas Ras o Rho ( GTPasas Ras o Rho ) y las fosfoinosítido 3-quinasas ( PI3K ). En los neutrófilos, los productos lipídicos de las PI3K regulan la activación de Rac1, Rac2 hematopoyético y RhoG GTPasas de la familia Rho y son necesarios para la motilidad celular . Las Ras-GTPasas y Rac-GTPasas regulan la dinámica del citoesqueleto y facilitan la adhesión, migración y propagación de los neutrófilos. [28] [29] [30] Se acumulan asimétricamente a la membrana plasmática en el borde delantero de las células polarizadas. Al regular espacialmente las GTPasas Rho y organizar el borde delantero de la célula, las PI3K y sus productos lipídicos podrían desempeñar un papel fundamental en el establecimiento de la polaridad de los leucocitos, como moléculas de brújula que le indican a la célula dónde arrastrarse.
Se ha demostrado en ratones que, en determinadas condiciones, los neutrófilos tienen un tipo específico de comportamiento migratorio denominado enjambre de neutrófilos , durante el cual migran de manera altamente coordinada y se acumulan y agrupan en sitios de inflamación. [31]
Función antimicrobiana
Al ser altamente móviles , los neutrófilos se congregan rápidamente en un foco de infección , atraídos por las citocinas expresadas por el endotelio activado , los mastocitos y los macrófagos . Los neutrófilos expresan [32] y liberan citocinas, que a su vez amplifican las reacciones inflamatorias de varios otros tipos de células.
Además de reclutar y activar otras células del sistema inmunológico, los neutrófilos desempeñan un papel clave en la defensa de primera línea contra los patógenos invasores y contienen una amplia gama de proteínas. [33] Los neutrófilos tienen tres métodos para atacar directamente a los microorganismos: fagocitosis (ingestión), desgranulación (liberación de antimicrobianos solubles) y generación de trampas extracelulares de neutrófilos (NET). [34]
Fagocitosis

Los neutrófilos son fagocitos capaces de ingerir microorganismos o partículas. Para que los reconozcan, deben estar recubiertos de opsoninas , un proceso conocido como opsonización de anticuerpos . [18] Pueden internalizar y matar muchos microbios , y cada evento fagocítico da como resultado la formación de un fagosoma en el que se secretan especies reactivas de oxígeno y enzimas hidrolíticas. El consumo de oxígeno durante la generación de especies reactivas de oxígeno se ha denominado " estallido respiratorio ", aunque no está relacionado con la respiración ni con la producción de energía.
El estallido respiratorio implica la activación de la enzima NADPH oxidasa , que produce grandes cantidades de superóxido , una especie reactiva del oxígeno. El superóxido se desintegra espontáneamente o se descompone a través de enzimas conocidas como superóxido dismutasas (Cu/ZnSOD y MnSOD), en peróxido de hidrógeno, que luego se convierte en ácido hipocloroso (HClO), por la enzima hemo verde mieloperoxidasa . Se cree que las propiedades bactericidas del HClO son suficientes para matar las bacterias fagocitadas por el neutrófilo, pero esto puede ser en cambio un paso necesario para la activación de las proteasas. [35]
Aunque los neutrófilos pueden matar a muchos microbios, la interacción de los neutrófilos con los microbios y las moléculas producidas por los microbios a menudo altera el recambio de los neutrófilos. La capacidad de los microbios para alterar el destino de los neutrófilos es muy variada, puede ser específica de cada microbio y va desde prolongar la vida útil de los neutrófilos hasta causar una lisis rápida de los neutrófilos después de la fagocitosis. Se ha informado que Chlamydia pneumoniae y Neisseria gonorrhoeae retrasan la apoptosis de los neutrófilos . [36] [37] [38] Por lo tanto, algunas bacterias, y aquellas que son patógenos predominantemente intracelulares, pueden extender la vida útil de los neutrófilos al interrumpir el proceso normal de apoptosis espontánea y/o PICD (muerte celular inducida por fagocitosis). En el otro extremo del espectro, algunos patógenos como Streptococcus pyogenes son capaces de alterar el destino de los neutrófilos después de la fagocitosis al promover una lisis celular rápida y/o acelerar la apoptosis hasta el punto de necrosis secundaria. [39] [40]
Desgranulación
Los neutrófilos también liberan una variedad de proteínas en tres tipos de gránulos mediante un proceso llamado desgranulación . El contenido de estos gránulos tiene propiedades antimicrobianas y ayuda a combatir las infecciones. Las células brillantes son neutrófilos leucocitos polimorfonucleares con gránulos. [41]
| Tipo de gránulo | Proteína |
|---|---|
| Gránulos azurófilos (o "gránulos primarios") | Mieloperoxidasa , proteína bactericida/aumentadora de la permeabilidad (BPI), defensinas y las serina proteasas elastasa de neutrófilos , proteinasa 3 y catepsina G |
| Gránulos específicos (o "gránulos secundarios") | Fosfatasa alcalina , lisozima , NADPH oxidasa , colagenasa , lactoferrina , histaminasa , [42] y catelicidina |
| Granulados terciarios | Catepsina , gelatinasa y colagenasa |
Trampas extracelulares de neutrófilos
En 2004, Brinkmann y sus colegas describieron una sorprendente observación de que la activación de los neutrófilos provoca la liberación de estructuras de ADN similares a redes; esto representa un tercer mecanismo para matar bacterias. [43] Estas trampas extracelulares de neutrófilos (NET) comprenden una red de fibras compuestas de cromatina y serina proteasas [44] que atrapan y matan a los microbios extracelulares. Se sugiere que las NET proporcionan una alta concentración local de componentes antimicrobianos y se unen, desarman y matan a los microbios independientemente de la captación fagocítica. Además de sus posibles propiedades antimicrobianas, las NET pueden servir como una barrera física que evita una mayor propagación de patógenos. La captura de bacterias puede ser un papel particularmente importante para las NET en la sepsis , donde las NET se forman dentro de los vasos sanguíneos. [45] Finalmente, se ha demostrado que la formación de NET aumenta la actividad bactericida de los macrófagos durante la infección. [46] [47] Recientemente, se ha demostrado que las NET desempeñan un papel en las enfermedades inflamatorias, ya que las NET podrían detectarse en la preeclampsia , un trastorno inflamatorio relacionado con el embarazo en el que se sabe que los neutrófilos se activan. [48] La formación de NET de neutrófilos también puede afectar la enfermedad cardiovascular , ya que las NET pueden influir en la formación de trombos en las arterias coronarias . [49] [50] Ahora se sabe que las NET exhiben efectos protrombóticos tanto in vitro [51] como in vivo . [52] [53] Más recientemente, en 2020, las NET estuvieron implicadas en la formación de coágulos sanguíneos en casos de COVID-19 grave . [54]
Neutrófilos asociados a tumores (TANS)
Los TAN pueden exhibir una tasa elevada de acidificación extracelular cuando hay un aumento en los niveles de glucólisis. [55] Cuando hay un cambio metabólico en los TAN, esto puede conducir a la progresión tumoral en ciertas áreas del cuerpo, como los pulmones. Los TAN apoyan el crecimiento y la progresión de los tumores a diferencia de los neutrófilos normales que inhibirían la progresión tumoral a través de la fagocitosis de las células tumorales. Utilizando un modelo de ratón, identificaron que tanto el metabolismo de Glut1 como el de la glucosa aumentaron en los TAN encontrados dentro de un ratón que poseía adenocarcinoma de pulmón. [55] Un estudio mostró que las células tumorales pulmonares pueden iniciar osteoblastos de forma remota y estos osteoblastos pueden empeorar los tumores de dos maneras. Primero, pueden inducir la formación de neutrófilos con alta expresión de SiglecF que a su vez promueve el crecimiento y la progresión del tumor pulmonar. Segundo, los osteoblastos pueden promover el crecimiento óseo formando así un entorno favorable para que las células tumorales crezcan para formar metástasis ósea. [56]
Importancia clínica

Los recuentos bajos de neutrófilos se denominan neutropenia . Puede ser congénita (desarrollada en el nacimiento o antes) o puede desarrollarse más tarde, como en el caso de la anemia aplásica o algunos tipos de leucemia . También puede ser un efecto secundario de la medicación , sobre todo la quimioterapia . La neutropenia hace que una persona sea muy susceptible a las infecciones. También puede ser el resultado de la colonización por parásitos neutrófilos intracelulares.
En la deficiencia de alfa 1-antitripsina , la importante elastasa de los neutrófilos no es inhibida adecuadamente por la alfa 1-antitripsina , lo que lleva a un daño tisular excesivo en presencia de inflamación, siendo la más destacada el enfisema . También se han demostrado efectos negativos de la elastasa en casos en los que los neutrófilos se activan excesivamente (en individuos por lo demás sanos) y liberan la enzima en el espacio extracelular. La actividad no regulada de la elastasa de los neutrófilos puede llevar a la alteración de la barrera pulmonar que muestra síntomas correspondientes a una lesión pulmonar aguda . [57] La enzima también influye en la actividad de los macrófagos al escindir sus receptores tipo toll (TLR) y regular a la baja la expresión de citocinas al inhibir la translocación nuclear de NF-κB . [58]
En la fiebre mediterránea familiar (FMF), una mutación en el gen de la pirina (o marenostrina ), que se expresa principalmente en los granulocitos neutrófilos, conduce a una respuesta de fase aguda constitutivamente activa y causa ataques de fiebre , artralgia , peritonitis y, eventualmente, amiloidosis . [59]
La hiperglucemia puede provocar disfunción de los neutrófilos. La disfunción de la vía bioquímica de los neutrófilos, la mieloperoxidasa , así como la reducción de la desgranulación, se asocian con la hiperglucemia. [60]
El recuento absoluto de neutrófilos (RAN) también se utiliza en el diagnóstico y pronóstico. El RAN es el estándar de oro para determinar la gravedad de la neutropenia y, por lo tanto, la fiebre neutropénica. Cualquier RAN < 1500 células / mm 3 se considera neutropenia, pero < 500 células / mm 3 se considera grave. [61] También hay nuevas investigaciones que vinculan el RAN con el infarto de miocardio como una ayuda en el diagnóstico temprano. [62] [63] Los neutrófilos promueven la taquicardia ventricular en el infarto agudo de miocardio. [64]
En la autopsia , la presencia de neutrófilos en el corazón o el cerebro es uno de los primeros signos de infarto y es útil para determinar el momento y el diagnóstico del infarto de miocardio y el accidente cerebrovascular .
- Los neutrófilos se observan en un infarto de miocardio aproximadamente a las 12-24 horas, [65] como se ve en esta micrografía .
- En el caso de un accidente cerebrovascular , comienzan a infiltrarse en el cerebro infartado después de 6 a 8 horas. [66]
![Los neutrófilos se observan en un infarto de miocardio aproximadamente entre las 12 y 24 horas,[65] como se ve en esta micrografía.](http://upload.wikimedia.org/wikipedia/commons/thumb/6/6c/Histopathology_of_neutrophil_infiltration_in_myocardial_infarction.jpg/1280px-Histopathology_of_neutrophil_infiltration_in_myocardial_infarction.jpg)
![En el caso de un accidente cerebrovascular, comienzan a infiltrarse en el cerebro infartado después de 6 a 8 horas.[66]](http://upload.wikimedia.org/wikipedia/commons/thumb/1/1e/Histopathology_of_thalamus_infarction_at_approximately_24_hours,_high_magnification,_annotated.jpg/1280px-Histopathology_of_thalamus_infarction_at_approximately_24_hours,_high_magnification,_annotated.jpg)
Evasión y resistencia de patógenos
Al igual que los fagocitos, los patógenos pueden evadir o infectar a los neutrófilos. [67] Algunos patógenos bacterianos desarrollaron diversos mecanismos, como moléculas de virulencia, para evitar ser eliminados por los neutrófilos. Estas moléculas en conjunto pueden alterar o interrumpir el reclutamiento de neutrófilos, la apoptosis o la actividad bactericida. [67]
Los neutrófilos también pueden servir como célula huésped para varios parásitos que los infectan evitando la fagocitosis, entre ellos:
- Leishmania major – utiliza neutrófilos como vehículo para parasitar fagocitos [26]
- M. tuberculosis [68]
- M. leprae [68]
- Yersinia pestis [68]
- Chlamydia pneumoniae [68]
Antígenos de neutrófilos
Hay cinco conjuntos (HNA 1–5) de antígenos de neutrófilos reconocidos. Los tres antígenos HNA-1 (ac) se encuentran en el receptor de baja afinidad Fc-γ IIIb (FCGR3B : CD16b ). El único antígeno conocido HNA-2a se encuentra en CD177 . El sistema de antígenos HNA-3 tiene dos antígenos (3a y 3b) que se encuentran en el séptimo exón del gen CLT2 ( SLC44A2 ). Los sistemas de antígenos HNA-4 y HNA-5 tienen cada uno dos antígenos conocidos (a y b) y se encuentran en la integrina β2 . HNA-4 se encuentra en la cadena αM ( CD11b ) y HNA-5 se encuentra en la unidad de integrina αL ( CD11a ). [69]
Subpoblaciones

Se identificaron dos subpoblaciones de neutrófilos funcionalmente desiguales en función de los diferentes niveles de generación de metabolitos reactivos de oxígeno, la permeabilidad de la membrana, la actividad del sistema enzimático y la capacidad de inactivación. Las células de una subpoblación con alta permeabilidad de la membrana (neutrófilos asesinos) generan intensamente metabolitos reactivos de oxígeno y se inactivan como consecuencia de la interacción con el sustrato, mientras que las células de otra subpoblación (neutrófilos enjaulados) producen especies reactivas de oxígeno con menor intensidad, no se adhieren al sustrato y conservan su actividad. [70] [71] [72] [73] [74] Estudios adicionales han demostrado que los tumores pulmonares pueden ser infiltrados por varias poblaciones de neutrófilos. [75]
Video
- Se puede observar un neutrófilo que se mueve rápidamente tomando varios conidios durante un tiempo de obtención de imágenes de 2 horas con un fotograma cada 30 segundos.
- Aquí se puede observar un neutrófilo que absorbe selectivamente varias levaduras Candida ( marcadas con fluorescencia en verde) a pesar de varios contactos con conidios de Aspergillus fumigatus (sin marcar, blanco/transparente) en una matriz de colágeno tridimensional . El tiempo de obtención de imágenes fue de 2 horas con un fotograma cada 30 segundos.
Los neutrófilos muestran una motilidad ameboide altamente direccional en las falanges y las almohadillas plantares infectadas. Se realizaron imágenes intravitales en la almohadilla plantar de ratones LysM-eGFP 20 minutos después de la infección con Listeria monocytogenes . [76]
Imágenes adicionales
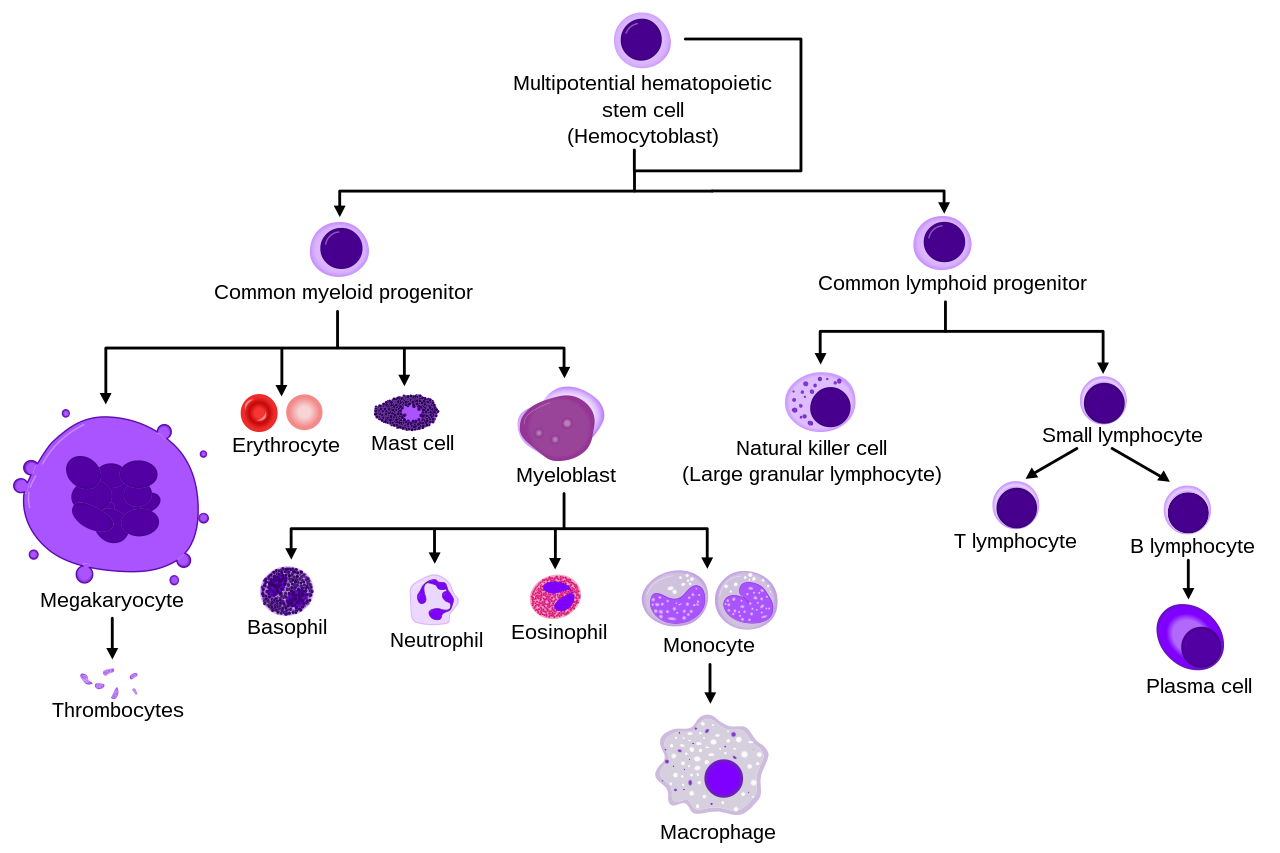
- Linaje de células sanguíneas
- Linajes más completos

_diagram_en.svg/1280px-Hematopoiesis_(human)_diagram_en.svg.png)
.png){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
_diagram_en.svg){kind=link}
Véase también
Referencias
- ^ Actor J (2012). Revisión integrada de Elsevier sobre inmunología y microbiología (segunda edición). doi :10.1016/B978-0-323-07447-6.00002-8.
- ^ Ermert D, Niemiec MJ, Röhm M, Glenthøj A, Borregaard N, Urban CF (agosto de 2013). "Candida albicans se escapa de los neutrófilos del ratón". Revista de biología de leucocitos . 94 (2): 223–236. doi :10.1189/jlb.0213063. PMID 23650619. S2CID 25619835.
- ^ Witko-Sarsat V, Rieu P, Descamps-Latscha B, Lesavre P, Halbwachs-Mecarelli L (mayo de 2000). "Neutrófilos: moléculas, funciones y aspectos fisiopatológicos". Investigación de laboratorio; una revista de métodos técnicos y patología . 80 (5): 617–653. doi : 10.1038/labinvest.3780067 . PMID 10830774. S2CID 22536645.
- ^ Klebanoff SJ, Clark RA (1978). El neutrófilo: función y trastornos clínicos . Elsevier/North-Holland Amsterdam. ISBN 978-0-444-80020-6.
- ^ Nathan C (marzo de 2006). "Neutrófilos e inmunidad: desafíos y oportunidades". Nature Reviews. Inmunología . 6 (3): 173–182. doi :10.1038/nri1785. PMID 16498448. S2CID 1590558.
- ^ Welsh CJ (2021). Fundamentos de anatomía y fisiología humana de Hole (14.ª ed.). Nueva York, EE. UU.: McGraw Hill. pág. 336. ISBN 978-1-260-57521-7. Recuperado el 28 de febrero de 2023 .
- ^ Jacobs L, Nawrot TS, de Geus B, Meeusen R, Degraeuwe B, Bernard A, et al. (octubre de 2010). "Respuestas subclínicas en ciclistas sanos expuestos brevemente a la contaminación del aire relacionada con el tráfico: un estudio de intervención". Salud ambiental . 9 (64): 64. Bibcode :2010EnvHe...9...64J. doi : 10.1186/1476-069X-9-64 . PMC 2984475 . PMID 20973949.
- ^ Waugh DJ, Wilson C (noviembre de 2008). "La vía de la interleucina-8 en el cáncer". Clinical Cancer Research . 14 (21): 6735–6741. doi :10.1158/1078-0432.CCR-07-4843. PMID 18980965. S2CID 9415085.
- ^ De Larco JE, Wuertz BR, Furcht LT (agosto de 2004). "El papel potencial de los neutrófilos en la promoción del fenotipo metastásico de tumores que liberan interleucina-8". Clinical Cancer Research . 10 (15): 4895–4900. doi :10.1158/1078-0432.CCR-03-0760. PMID 15297389. S2CID 9782495.
- ^ Yoo SK, Starnes TW, Deng Q, Huttenlocher A (noviembre de 2011). "Lyn es un sensor redox que media la atracción de leucocitos por heridas in vivo". Nature . 480 (7375): 109–112. Bibcode :2011Natur.480..109Y. doi :10.1038/nature10632. PMC 3228893 . PMID 22101434.
- ^ Barer MR (2012). "La historia natural de la infección". Microbiología médica . Elsevier. págs. 168-173. doi :10.1016/b978-0-7020-4089-4.00029-9. ISBN 978-0-7020-4089-4.
- ^ Cohen S, Burns RC (2002). Caminos de la pulpa (8.ª ed.). St. Louis: Mosby. pág. 465.
- ^ Huerta MÁ, Molina-Álvarez M, García MM, Tejada MA, Goicoechea C, Ghasemlou N, et al. (2024-10-25). "El papel de los neutrófilos en el dolor: revisión sistemática y metaanálisis de estudios en animales". Pain . doi :10.1097/j.pain.0000000000003450. ISSN 0304-3959.
- ^ Niemiec MJ, De Samber B, Garrevoet J, Vergucht E, Vekemans B, De Rycke R, et al. (junio de 2015). "Paisaje de oligoelementos de neutrófilos humanos activados y en reposo a nivel submicrométrico". Metalómica . 7 (6): 996–1010. doi : 10.1039/c4mt00346b . PMID 25832493.
- ^ abc Zucker-Franklin D, Greaves MF, Grossi CE, Marmont AM (1988). "Neutrófilos". Atlas de células sanguíneas: función y patología . Vol. 1 (2.ª ed.). Filadelfia: Lea & Ferbiger. ISBN 978-0-8121-1094-4.
- ^ Karni RJ, Wangh LJ, Sanchez JA (agosto de 2001). "Ubicación y orientación no aleatorias del cromosoma X inactivo en los núcleos de neutrófilos humanos". Chromosoma . 110 (4): 267–274. doi :10.1007/s004120100145. PMID 11534818. S2CID 24750407.
- ^ Reich D, Nalls MA, Kao WH, Akylbekova EL, Tandon A, Patterson N, et al. (enero de 2009). "El recuento reducido de neutrófilos en personas de ascendencia africana se debe a una variante reguladora en el gen del receptor de antígeno Duffy para quimiocinas". PLOS Genetics . 5 (1): e1000360. doi : 10.1371/journal.pgen.1000360 . PMC 2628742 . PMID 19180233.
- ^ ab Edwards SW (1994). Bioquímica y fisiología de los neutrófilos . Cambridge University Press. pág. 6. ISBN 978-0-521-41698-6.
- ^ Sanchez A, Reeser JL, Lau HS, Yahiku PY, Willard RE, McMillan PJ, et al. (noviembre de 1973). "Role of sugars in human neutrophilic phagocytosis". The American Journal of Clinical Nutrition . 26 (11): 1180–1184. doi : 10.1093/ajcn/26.11.1180 . PMID 4748178.
Estos datos sugieren que la función y no el número de fagocitos se alteró por la ingestión de azúcares. Esto implica a la glucosa y otros carbohidratos simples en el control de la fagocitosis y muestra que los efectos duran al menos 5 horas. Por otro lado, un ayuno de 36 o 60 horas aumentó significativamente (P < 0,001) el índice fagocítico.
- ^ Rubin-Bejerano I, Abeijon C, Magnelli P, Grisafi P, Fink GR (julio de 2007). "La fagocitosis por neutrófilos humanos es estimulada por un componente único de la pared celular fúngica". Cell Host & Microbe . 2 (1): 55–67. doi :10.1016/j.chom.2007.06.002. PMC 2083279 . PMID 18005717.
- ^ Kneller A (2007). "Los glóbulos blancos son exigentes con el azúcar". Instituto Whitehead . Consultado el 9 de agosto de 2013 .
- ^ Tak T, Tesselaar K, Pillay J, Borghans JA, Koenderman L (octubre de 2013). "¿Cuál es tu edad otra vez? Se revisó la determinación de las semividas de los neutrófilos humanos". Journal of Leukocyte Biology . 94 (4): 595–601. doi :10.1189/jlb.1112571. PMID 23625199. S2CID 40113921.
- ^ Pillay J, den Braber I, Vrisekoop N, Kwast LM, de Boer RJ, Borghans JA, et al. (julio de 2010). "El marcado in vivo con 2H2O revela una vida útil de los neutrófilos humanos de 5,4 días". Blood . 116 (4): 625–627. doi : 10.1182/blood-2010-01-259028 . PMID 20410504. S2CID 909519.
- ^ ab Wheater PR, Stevens A (2002). Histopatología básica de Wheater: un atlas en color y texto . Edimburgo: Churchill Livingstone. ISBN 978-0-443-07001-3.
- ^ Akbar N, Braithwaite AT, Corr EM, Koelwyn GJ, van Solingen C, Cochain C, et al. (marzo de 2023). "Movilización rápida de neutrófilos por vesículas extracelulares derivadas de células endoteliales VCAM-1+". Investigación cardiovascular . 119 (1): 236–251. doi :10.1093/cvr/cvac012. PMC 10022859 . PMID 35134856.
- ^ ab Ritter U, Frischknecht F, van Zandbergen G (noviembre de 2009). "¿Son los neutrófilos células hospedadoras importantes para los parásitos de Leishmania?". Trends in Parasitology . 25 (11): 505–510. doi :10.1016/j.pt.2009.08.003. PMID 19762280.
- ^ Serhan CN, Ward PA, Gilroy DW (2010). Fundamentos de la inflamación. Cambridge University Press. págs. 53-54. ISBN 978-0-521-88729-8.
- ^ Pantarelli C, Welch HC (noviembre de 2018). "Rac-GTPasas y Rac-GEF en la adhesión, migración y reclutamiento de neutrófilos". Revista Europea de Investigación Clínica . 48 (Supl. 2): e12939. doi :10.1111/eci.12939. PMC 6321979 . PMID 29682742.
- ^ Lin Y, Pal DS, Banerjee P, Banerjee T, Qin G, Deng Y, et al. (julio de 2024). "La supresión de Ras potencia la polarización y migración celular impulsada por la contractilidad de actomiosina trasera". Nature Cell Biology : 1–15. doi :10.1038/s41556-024-01453-4. PMID 38951708.
- ^ Pal DS, Banerjee T, Lin Y, de Trogoff F, Borleis J, Iglesias PA, et al. (julio de 2023). "La activación de nodos descendentes individuales en la red de factores de crecimiento dirige la migración de células inmunitarias". Developmental Cell . 58 (13): 1170–1188.e7. doi :10.1016/j.devcel.2023.04.019. PMC 10524337 . PMID 37220748.
- ^ Lämmermann T, Afonso PV, Angermann BR, Wang JM, Kastenmüller W, Parent CA, et al. (junio de 2013). "Los enjambres de neutrófilos requieren LTB4 e integrinas en los sitios de muerte celular in vivo". Nature . 498 (7454): 371–375. Bibcode :2013Natur.498..371L. doi :10.1038/nature12175. PMC 3879961 . PMID 23708969.
- ^ Ear T, McDonald PP (abril de 2008). "Generación de citocinas, activación del promotor y activación de NF-kappaB independiente del oxidante en un modelo celular neutrofílico humano transfectable". BMC Immunology . 9 : 14. doi : 10.1186/1471-2172-9-14 . PMC 2322942 . PMID 18405381.
- ^ Ambatipudi KS, Old JM, Guilhaus M, Raftery M, Hinds L, Deane EM (2006). Análisis proteómico de las proteínas de los neutrófilos del ualabí de Tammar ( Macropus eugenii ). Comparative Biochemistry and Physiology. Parte D: Genómica y proteómica. 1(3), 283-291. DOI: 10.1016/j.cbd.2006.05.002
- ^ Hickey MJ, Kubes P (mayo de 2009). "Inmunidad intravascular: el encuentro huésped-patógeno en los vasos sanguíneos". Nature Reviews. Inmunología . 9 (5): 364–375. doi :10.1038/nri2532. PMID 19390567. S2CID 8068543.
- ^ Segal AW (2005). "Cómo los neutrófilos matan a los microbios". Revisión anual de inmunología . 23 (5): 197–223. doi :10.1146/annurev.immunol.23.021704.115653. PMC 2092448 . PMID 15771570.
- ^ Simons MP, Nauseef WM, Griffith TS, Apicella MA (noviembre de 2006). "Neisseria gonorrhoeae retrasa el inicio de la apoptosis en leucocitos polimorfonucleares". Microbiología celular . 8 (11): 1780–1790. doi :10.1111/j.1462-5822.2006.00748.x. PMID 16803582. S2CID 25253422.
- ^ Chen A, Seifert HS (noviembre de 2011). "Inhibición de la señalización apoptótica en leucocitos polimorfonucleares mediada por Neisseria gonorrhoeae". Infección e inmunidad . 79 (11): 4447–4458. doi :10.1128/IAI.01267-10. PMC 3257915 . PMID 21844239.
- ^ van Zandbergen G, Gieffers J, Kothe H, Rupp J, Bollinger A, Aga E, et al. (febrero de 2004). "Chlamydia pneumoniae se multiplica en los granulocitos neutrófilos y retrasa su apoptosis espontánea". Journal of Immunology . 172 (3): 1768–1776. doi : 10.4049/jimmunol.172.3.1768 . PMID 14734760. S2CID 27422510.
- ^ Kobayashi SD, Braughton KR, Whitney AR, Voyich JM, Schwan TG, Musser JM, et al. (septiembre de 2003). "Los patógenos bacterianos modulan un programa de diferenciación por apoptosis en neutrófilos humanos". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 100 (19): 10948–10953. doi : 10.1073/pnas.1833375100 . PMC 196908 . PMID 12960399.
- ^ Kobayashi SD, Malachowa N, DeLeo FR (2017). "Influencia de los microbios en la vida y muerte de los neutrófilos". Frontiers in Cellular and Infection Microbiology . 7 (159): 159. doi : 10.3389/fcimb.2017.00159 . PMC 5410578 . PMID 28507953.
 El material fue copiado de esta fuente, que está disponible bajo una Licencia Creative Commons Atribución 4.0 Internacional.
El material fue copiado de esta fuente, que está disponible bajo una Licencia Creative Commons Atribución 4.0 Internacional. - ^ Berman LB, Feys JO, Schreiner GE (noviembre de 1956). "Observaciones sobre el fenómeno de las células brillantes". The New England Journal of Medicine . 255 (21): 989–991. doi :10.1056/NEJM195611222552104. PMID 13378597.
- ^ Ringel EW, Soter NA, Austen KF (agosto de 1984). "Localización de la histaminasa en el gránulo específico del neutrófilo humano". Inmunología . 52 (4): 649–658. PMC 1454675 . PMID 6430792.
- ^ Brinkmann V, Reichard U, Goosmann C, Fauler B, Uhlemann Y, Weiss DS, et al. (marzo de 2004). "Las trampas extracelulares de neutrófilos matan bacterias". Science . 303 (5663): 1532–1535. Bibcode :2004Sci...303.1532B. doi :10.1126/science.1092385. PMID 15001782. S2CID 21628300.
- ^ Urban CF, Ermert D, Schmid M, Abu-Abed U, Goosmann C, Nacken W, et al. (octubre de 2009). "Las trampas extracelulares de neutrófilos contienen calprotectina, un complejo proteico citosólico implicado en la defensa del huésped contra Candida albicans". PLOS Pathogens . 5 (10): e1000639. doi : 10.1371/journal.ppat.1000639 . PMC 2763347 . PMID 19876394.
- ^ Clark SR, Ma AC, Tavener SA, McDonald B, Goodarzi Z, Kelly MM, et al. (abril de 2007). "El TLR4 plaquetario activa las trampas extracelulares de neutrófilos para atrapar bacterias en sangre séptica". Nature Medicine . 13 (4): 463–469. doi :10.1038/nm1565. PMID 17384648. S2CID 22372863.
- ^ Monteith AJ, Miller JM, Maxwell CN, Chazin WJ, Skaar EP (septiembre de 2021). "Las trampas extracelulares de neutrófilos mejoran la eliminación de patógenos bacterianos por parte de los macrófagos". Science Advances . 7 (37): eabj2101. Bibcode :2021SciA....7.2101M. doi :10.1126/sciadv.abj2101. PMC 8442908 . PMID 34516771.
- ^ Monteith AJ, Miller JM, Beavers WN, Maloney KN, Seifert EL, Hajnoczky G, et al. (febrero de 2022). "El uniportador de calcio mitocondrial afecta la actividad bactericida de los neutrófilos durante la infección por Staphylococcus aureus". Infección e inmunidad . 90 (2): e0055121. doi :10.1128/IAI.00551-21. PMC 8853686 . PMID 34871043.
- ^ Gupta AK, Hasler P, Holzgreve W, Hahn S (junio de 2007). "NET de neutrófilos: ¿un nuevo contribuyente a la hipoxia placentaria asociada a la preeclampsia?". Seminarios en inmunopatología . 29 (2): 163–167. doi :10.1007/s00281-007-0073-4. PMID 17621701. S2CID 12887059.
- ^ Hoyer FF, Nahrendorf M (febrero de 2017). "Contribuciones de los neutrófilos a la cardiopatía isquémica". European Heart Journal . 38 (7): 465–472. doi : 10.1093/eurheartj/ehx017 . PMID 28363210.
- ^ Mangold A, Alias S, Scherz T, Hofbauer M, Jakowitsch J, Panzenböck A, et al. (marzo de 2015). "La carga de trampa extracelular de neutrófilos coronarios y la actividad de la desoxirribonucleasa en el síndrome coronario agudo con elevación del segmento ST son predictores de la resolución del segmento ST y del tamaño del infarto". Circulation Research . 116 (7): 1182–1192. doi : 10.1161/CIRCRESAHA.116.304944 . PMID 25547404. S2CID 2532741.
- ^ Fuchs TA, Brill A, Duerschmied D, Schatzberg D, Monestier M, Myers DD, et al. (septiembre de 2010). "Las trampas de ADN extracelular promueven la trombosis". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 107 (36): 15880–15885. Bibcode :2010PNAS..10715880F. doi : 10.1073/pnas.1005743107 . PMC 2936604 . PMID 20798043.
- ^ Brill A, Fuchs TA, Savchenko AS, Thomas GM, Martinod K, De Meyer SF, et al. (enero de 2012). "Las trampas extracelulares de neutrófilos promueven la trombosis venosa profunda en ratones". Journal of Thrombosis and Haemostasis . 10 (1): 136–144. doi :10.1111/j.1538-7836.2011.04544.x. PMC 3319651 . PMID 22044575.
- ^ Borissoff JI, ten Cate H (septiembre de 2011). "Desde la liberación de trampas extracelulares de neutrófilos hasta la trombosis: ¿un mecanismo de defensa del huésped que se dispara?". Journal of Thrombosis and Haemostasis . 9 (9): 1791–1794. doi : 10.1111/j.1538-7836.2011.04425.x . PMID 21718435. S2CID 5368241.
- ^ Zuo Y, Yalavarthi S, Shi H, Gockman K, Zuo M, Madison JA, et al. (junio de 2020). "Trampas extracelulares de neutrófilos en COVID-19". Perspectiva de la JCI . 5 (11): e138999. doi : 10.1172/jci.insight.138999. PMC 7308057 . PMID 32329756. S2CID 216109364.
- ^ ab Ancey PB, Contat C, Boivin G, Sabatino S, Pascual J, Zangger N, et al. (mayo de 2021). "La expresión de GLUT1 en neutrófilos asociados a tumores promueve el crecimiento del cáncer de pulmón y la resistencia a la radioterapia". Investigación sobre el cáncer . 81 (9): 2345–2357. doi :10.1158/0008-5472.CAN-20-2870. PMC 8137580 . PMID 33753374.
- ^ Azevedo PO, Paiva AE, Santos GS, Lousado L, Andreotti JP, Sena IF, et al. (diciembre de 2018). "La comunicación cruzada entre el cáncer de pulmón y los huesos da como resultado neutrófilos que promueven la progresión tumoral". Cancer and Metastasis Reviews . 37 (4): 779–790. doi :10.1007/s10555-018-9759-4. PMC 6358512 . PMID 30203108.
- ^ Kawabata K, Hagio T, Matsuoka S (septiembre de 2002). "El papel de la elastasa de neutrófilos en la lesión pulmonar aguda". Revista Europea de Farmacología . 451 (1): 1–10. doi :10.1016/S0014-2999(02)02182-9. PMID 12223222.
- ^ Domon H, Nagai K, Maekawa T, Oda M, Yonezawa D, Takeda W, et al. (2018). "La elastasa de neutrófilos subvierte la respuesta inmunitaria al escindir los receptores tipo Toll y las citocinas en la neumonía neumocócica". Frontiers in Immunology . 9 : 732. doi : 10.3389/fimmu.2018.00732 . PMC 5996908 . PMID 29922273.
- ^ Ozen S (julio de 2003). "Fiebre mediterránea familiar: revisitando una enfermedad antigua". Revista Europea de Pediatría . 162 (7–8): 449–454. doi :10.1007/s00431-003-1223-x. PMID 12751000. S2CID 3464945.
- ^ Xiu F, Stanojcic M, Diao L, Jeschke MG (8 de mayo de 2014). "Hiperglucemia por estrés, tratamiento con insulina y células inmunes innatas". Revista Internacional de Endocrinología . 2014 : 486403. doi : 10.1155/2014/486403 . PMC 4034653. PMID 24899891 .
- ^ Al-Gwaiz LA, Babay HH (2007). "El valor diagnóstico del recuento absoluto de neutrófilos, el recuento de bandas y los cambios morfológicos de los neutrófilos en la predicción de infecciones bacterianas". Principios y práctica médica . 16 (5): 344–347. doi : 10.1159/000104806 . PMID 17709921. S2CID 5499290.
- ^ Khan HA, Alhomida AS, Sobki SH, Moghairi AA, Koronki HE (2012). "Recuento de células sanguíneas y su correlación con la creatina quinasa y la proteína C reactiva en pacientes con infarto agudo de miocardio". Revista internacional de medicina clínica y experimental . 5 (1): 50–55. PMC 3272686 . PMID 22328948.
- ^ Basili S, Di Francoi M, Rosa A, Ferroni P, Diurni V, Scarpellini MG y col. (Abril de 2004). "Recuento absoluto de neutrófilos y niveles de fibrinógeno como ayuda en el diagnóstico precoz del infarto agudo de miocardio". Acta Cardiológica . 59 (2): 135-140. doi :10.2143/ac.59.2.2005167. PMID 15139653. S2CID 37382677.
- ^ Grune J, Lewis AJ, Yamazoe M, Hulsmans M, Rohde D, Xiao L, et al. (julio de 2022). "Los neutrófilos incitan y los macrófagos evitan la tormenta eléctrica después de un infarto de miocardio". Nature Cardiovascular Research . 1 (7): 649–664. doi :10.1038/s44161-022-00094-w. PMC 9410341 . PMID 36034743.
- ^ Michaud K, Basso C, d'Amati G, Giordano C, Kholová I, Preston SD, et al. (febrero de 2020). "Diagnóstico de infarto de miocardio en la autopsia: reevaluación de la AECVP a la luz de la clasificación clínica actual". Virchows Archiv . 476 (2): 179–194. doi : 10.1007/s00428-019-02662-1 . PMC: 7028821. PMID: 31522288 .
"Este artículo se distribuye bajo los términos de la Licencia Creative Commons Atribución 4.0 Internacional (http://creativecommons.org/licenses/by/4.0/ Archivado el 21 de noviembre de 2015 en Wayback Machine )" - ^ Jickling GC, Liu D, Ander BP, Stamova B, Zhan X, Sharp FR (junio de 2015). "Ataque a los neutrófilos en el accidente cerebrovascular isquémico: perspectivas translacionales de estudios experimentales". Journal of Cerebral Blood Flow and Metabolism . 35 (6): 888–901. doi :10.1038/jcbfm.2015.45. PMC 4640255 . PMID 25806703.
- ^ ab Kobayashi SD, Malachowa N, DeLeo FR (2018). "Neutrófilos y evasión inmunitaria bacteriana". Revista de inmunidad innata . 10 (5–6): 432–441. doi :10.1159/000487756. PMC 6784029 . PMID 29642066.
- ^ abcd Parker HA, Forrester L, Kaldor CD, Dickerhof N, Hampton MB (23 de diciembre de 2021). "Actividad antimicrobiana de los neutrófilos contra las micobacterias". Frontiers in Immunology . 12 : 782495. doi : 10.3389/fimmu.2021.782495 . PMC 8732375 . PMID 35003097.
- ^ Chu HT, Lin H, Tsao TT, Chang CF, Hsiao WW, Yeh TJ, et al. (septiembre de 2013). "Genotipificación de antígenos de neutrófilos humanos (HNA) a partir de datos de secuenciación del genoma completo". BMC Medical Genomics . 6 (1): 31. doi : 10.1186/1755-8794-6-31 . PMC 3849977 . PMID 24028078. Este artículo incorpora texto disponible bajo la licencia CC BY 2.0.
- ^ ab Ignatov DY (2012). Heterogeneidad funcional de los neutrófilos humanos y su papel en la regulación de la cantidad de leucocitos en sangre periférica (PhD). Universidad Médica Nacional de Donetsk. doi :10.13140/RG.2.2.35542.34884.
- ^ Gerasimov IG, Ignatov DI (2001). "[Heterogenicidad funcional de los neutrófilos de la sangre humana: generación de especies activas de oxígeno]". Tsitologiia . 43 (5): 432–436. PMID 11517658.
- ^ Gerasimov IG, Ignatov DI (2004). "[Activación de neutrófilos in vitro]". Tsitología . 46 (2): 155-158. PMID 15174354.
- ^ Gerasimov IG, Ignatov DI, Kotel'nitskiĭ MA (2005). "[Reducción de tetrazolio azul de nitro por neutrófilos de sangre humana. I. La influencia del pH]". Tsitologiia . 47 (6): 549–553. PMID 16708848.
- ^ Gerasimov IG, Ignatov DI (2005). "[Reducción de tetrazolio azul de nitro por neutrófilos de sangre humana. II. La influencia de los iones de sodio y potasio]". Tsitologiia . 47 (6): 554–558. PMID 16708849.
- ^ Zilionis R, Engblom C, Pfirschke C, Savova V, Zemmour D, Saatcioglu HD, et al. (mayo de 2019). "La transcriptómica unicelular de cánceres de pulmón humanos y de ratón revela poblaciones mieloides conservadas en individuos y especies". Inmunidad . 50 (5): 1317–1334.e10. doi :10.1016/j.immuni.2019.03.009. PMC 6620049 . PMID 30979687.
- ^ Graham DB, Zinselmeyer BH, Mascarenhas F, Delgado R, Miller MJ, Swat W (2009). Unutmaz D (ed.). "La señalización de ITAM por factores de intercambio de nucleótidos de guanina Rho de la familia Vav regula las tasas de tránsito intersticial de los neutrófilos in vivo". PLOS ONE . 4 (2): e4652. Bibcode :2009PLoSO...4.4652G. doi : 10.1371/journal.pone.0004652 . PMC 2645696 . PMID 19247495.
{kind=link}
Enlaces externos
- Información sobre la neutropenia Archivado el 2 de diciembre de 2015 en Wayback Machine
- Calculadora del recuento absoluto de neutrófilos
- Contenido y distribución de oligoelementos en los neutrófilos