Ctenóforos
| Medusas peine | |
|---|---|
 | |
| "Ctenophorae" (medusa peine) | |
| Clasificación científica | |
| Dominio: | Eucariota |
| Reino: | Animalia |
| Filo: | Ctenófora de Eschscholtz , 1829 |
| Clases | |
{kind=link}
Ctenophora ( / t ə ˈ n ɒ f ər ə / tə- NOF -ər-ə ; sg. : ctenóforo / ˈ t ɛ n ə f ɔːr , ˈ t iː n ə -/ TEN -ə-for, TEE -nə- ; del griego antiguo κτείς (kteis) 'peine' y φέρω (pherō) 'llevar') [6] comprende un filo de invertebrados marinos , comúnmente conocidos como medusas peine , que habitan las aguas marinas de todo el mundo. Son notables por los grupos de cilios que utilizan para nadar (comúnmente denominados "peines"), y son los animales más grandes que nadan con la ayuda de cilios.
Dependiendo de la especie, los ctenóforos adultos miden desde unos pocos milímetros hasta 1,5 metros. Actualmente se reconocen solo 186 especies vivas. [7]
Sus cuerpos consisten en una masa de gelatina, con una capa de dos células de espesor en el exterior, y otra que recubre la cavidad interna. El filo tiene una amplia gama de formas corporales, incluyendo los cídipidos con forma de huevo con un par de tentáculos retráctiles que capturan presas, los platicténidos planos, generalmente sin peine, y los beroideos de boca grande , que se alimentan de otros ctenóforos.
Casi todos los ctenóforos funcionan como depredadores , capturando presas que van desde larvas microscópicas y rotíferos hasta adultos de pequeños crustáceos ; las excepciones son los juveniles de dos especies, que viven como parásitos de las salpas de las que se alimentan los adultos de su especie.
A pesar de sus cuerpos blandos y gelatinosos, los fósiles que se cree que representan ctenóforos aparecen en Lagerstätten y datan del Cámbrico temprano , hace unos 525 millones de años. La posición de los ctenóforos en el "árbol de la vida" ha sido debatida durante mucho tiempo en estudios de filogenética molecular. Los biólogos propusieron que los ctenóforos constituyen el segundo linaje animal ramificado más antiguo, siendo las esponjas el grupo hermano de todos los demás animales multicelulares ( hipótesis de la hermana Porifera ). [8] Otros biólogos sostienen que los ctenóforos surgieron antes que las esponjas ( hipótesis de la hermana Ctenophora ), que a su vez aparecieron antes de la división entre cnidarios y bilaterales . [9] [10] Pisani et al . volvieron a analizar los datos y sugirieron que los algoritmos informáticos utilizados para el análisis fueron engañados por la presencia de genes específicos de ctenóforos que eran marcadamente diferentes de los de otras especies. [11] [12] [ página necesaria ] Análisis de seguimiento de Whelan et al. (2017) [13] brindó mayor apoyo a la hipótesis de la "hermana de Ctenophora"; la cuestión sigue siendo un tema de disputa taxonómica. [14] [15] Schultz et al. (2023) encontraron cambios irreversibles en la sintenia en la hermana de Ctenophora, Myriazoa , que consiste en el resto de los animales. [16]
Características distintivas
{kind=link}

a Beroe ovata , b cídípido no identificado, c cídípido "Tortugas rojo",
d Bathocyroe fosteri , e Mnemiopsis leidyi , y f Ocyropsis sp. [17]
Entre los filos animales, los ctenóforos son más complejos que las esponjas , casi tan complejos como los cnidarios ( medusas , anémonas de mar , etc.) y menos complejos que los bilaterales (que incluyen a casi todos los demás animales). A diferencia de las esponjas, tanto los ctenóforos como los cnidarios tienen:
- células unidas por conexiones intercelulares y
- membranas basales tipo alfombra ;
- músculos ;
- sistemas nerviosos ; y
- órganos sensoriales (en algunos, no en todos).
Los ctenóforos se distinguen de todos los demás animales por tener coloblastos , que son pegajosos y se adhieren a las presas, aunque algunas especies de ctenóforos carecen de ellos. [18] [19]
Al igual que los cnidarios, los ctenóforos tienen dos capas principales de células que intercalan una capa intermedia de material gelatinoso, que se llama mesoglea en cnidarios y ctenóforos; los animales más complejos tienen tres capas celulares principales y ninguna capa intermedia gelatinosa. Por lo tanto, los ctenóforos y los cnidarios han sido etiquetados tradicionalmente como diploblásticos . [18] [20] Tanto los ctenóforos como los cnidarios tienen un tipo de músculo que, en animales más complejos, surge de la capa celular media , [21] y, como resultado, algunos libros de texto recientes clasifican a los ctenóforos como triploblásticos , [22] mientras que otros todavía los consideran diploblásticos. [18] Las medusas peine tienen más de 80 tipos de células diferentes , superando los números de otros grupos como placozoos, esponjas, cnidarios y algunos bilaterales de ramificación profunda. [23]
Los ctenóforos , que miden entre 1 milímetro (0,04 pulgadas) y 1,5 metros (5 pies), [22] [24] son los animales no coloniales más grandes que utilizan cilios ("pelos") como su principal método de locomoción. [22] La mayoría de las especies tienen ocho tiras, llamadas filas de peines, que recorren la longitud de sus cuerpos y llevan bandas de cilios similares a peines, llamadas "ctenes", apiladas a lo largo de las filas de peines de modo que cuando los cilios se agitan, los de cada peine tocan el peine de abajo. [22] El nombre "ctenophora" significa "que lleva peines", del griego κτείς (forma de tallo κτεν- ) que significa "peine" y el sufijo griego -φορος que significa "que lleva". [25]
| Esponjas [26] [27] | Cnidarios [18] [20] [28] | Ctenóforos [18] [22] | Bilateral [18] | |
|---|---|---|---|---|
| Cnidocitos | No | Sí | Sólo en algunas especies (obtenidas de cnidarios ingeridos) | |
| microARN | Sí | Sí | No | Sí |
| Genes Hox | No | Sí | No | Sí |
| Coloblastos | No | En la mayoría de las especies [19] | No | |
| Órganos digestivos y circulatorios | No | Sí | ||
| Poros anales | No | Sí | Mayormente sí | |
| Número de capas principales de células | Dos, con una capa gelatinosa entre ellos. | Debate sobre si dos [18] o tres [21] [22] | Tres | |
| Las células de cada capa están unidas entre sí | No, excepto que los Homoscleromorpha tienen membranas basales [29] | Sí: Conexiones intercelulares; membranas basales | ||
| Órganos sensoriales | No | Sí | ||
| Ojos (por ejemplo, ocelos ) | No | Sí | No | Sí |
| Órgano apical | No | Sí | Sí | En especies con larvas primarias ciliadas |
| Abundancia de células en la capa intermedia de "gelatina" | Muchos | Pocos | [no aplicable] | |
| Las células de la capa externa pueden moverse hacia adentro y cambiar sus funciones. | Sí | No | ||
| Sistema nervioso | No | Sí, sencillo. | De lo simple a lo complejo | |
| Musculatura | Ninguno | Mayormente epiteliomuscular | Mayormente mioepitelial | Principalmente miocitos |
Descripción
Para un filo con relativamente pocas especies, los ctenóforos tienen una amplia gama de planes corporales. [22] Las especies costeras deben ser lo suficientemente resistentes para soportar olas y partículas de sedimento arremolinadas, mientras que algunas especies oceánicas son tan frágiles que es muy difícil capturarlas intactas para su estudio. [19] Además, las especies oceánicas no se conservan bien, [19] y se conocen principalmente a partir de fotografías y notas de observadores. [30] Por lo tanto, hasta hace poco la mayor parte de la atención se ha concentrado en tres géneros costeros : Pleurobrachia , Beroe y Mnemiopsis . [19] [31] Al menos dos libros de texto basan sus descripciones de los ctenóforos en el cídipido Pleurobrachia . [18] [22]
Dado que el cuerpo de muchas especies es casi radialmente simétrico , el eje principal es oral a aboral (desde la boca hasta el extremo opuesto). Sin embargo, dado que solo dos de los canales cerca del estatocisto terminan en poros anales , los ctenóforos no tienen simetría especular, aunque muchos tienen simetría rotacional. En otras palabras, si el animal gira en un semicírculo, se ve igual que cuando comenzó. [32]
Características comunes
El filo Ctenóforo tiene una amplia gama de formas corporales, incluidos los platicténidos aplanados de aguas profundas , en los que los adultos de la mayoría de las especies carecen de peines, y los beroideos costeros, que carecen de tentáculos y se aprovechan de otros ctenóforos utilizando enormes bocas armadas con grupos de cilios grandes y rígidos que actúan como dientes.
Capas del cuerpo
{kind=link}

Al igual que los de los cnidarios ( medusas , anémonas de mar , etc.), los cuerpos de los ctenóforos consisten en una mesoglea relativamente gruesa y gelatinosa intercalada entre dos epitelios , capas de células unidas por conexiones intercelulares y por una membrana basal fibrosa que secretan . [18] [22] Los epitelios de los ctenóforos tienen dos capas de células en lugar de una, y algunas de las células de la capa superior tienen varios cilios por célula. [22]
La capa externa de la epidermis (piel exterior) consta de: células sensoriales; células que secretan moco , que protege el cuerpo; y células intersticiales, que pueden transformarse en otros tipos de células. En partes especializadas del cuerpo, la capa externa también contiene coloblastos , que se encuentran a lo largo de la superficie de los tentáculos y se utilizan para capturar presas, o células que tienen múltiples cilios grandes, para la locomoción. La capa interna de la epidermis contiene una red nerviosa y células mioepiteliales que actúan como músculos . [22]
La cavidad interna forma: una boca que generalmente se puede cerrar mediante músculos; una faringe ("garganta"); un área más amplia en el centro que actúa como estómago ; y un sistema de canales internos. Estos se ramifican a través de la mesoglea hacia las partes más activas del animal: la boca y la faringe; las raíces de los tentáculos, si están presentes; a lo largo de la parte inferior de cada fila de peines; y cuatro ramas alrededor del complejo sensorial en el extremo más alejado de la boca; dos de estas cuatro ramas terminan en poros anales . La superficie interna de la cavidad está revestida con un epitelio , la gastrodermis . La boca y la faringe tienen cilios y músculos bien desarrollados. En otras partes del sistema de canales, la gastrodermis es diferente en los lados más cercanos y más alejados del órgano que irriga. El lado más cercano está compuesto por células nutritivas altas que almacenan nutrientes en vacuolas (compartimentos internos), células germinales que producen óvulos o espermatozoides y fotocitos que producen bioluminiscencia . El lado más alejado del órgano está cubierto de células ciliadas que hacen circular agua a través de los canales, puntuados por rosetas ciliares, poros que están rodeados por dobles verticilos de cilios y se conectan a la mesoglea. [22]
Alimentación, excreción y respiración.
Cuando se traga una presa, se licua en la faringe mediante enzimas y contracciones musculares de la faringe. La suspensión resultante se transporta a través del sistema de canales mediante el movimiento de los cilios y es digerida por las células nutritivas. Las rosetas ciliares de los canales pueden ayudar a transportar nutrientes a los músculos de la mesoglea. Los poros anales pueden expulsar pequeñas partículas no deseadas, pero la mayor parte de la materia no deseada se regurgita a través de la boca. [22]
Se sabe poco sobre cómo los ctenóforos se deshacen de los desechos producidos por las células. Las rosetas ciliares de la gastrodermis pueden ayudar a eliminar los desechos de la mesoglea y también pueden ayudar a ajustar la flotabilidad del animal bombeando agua hacia dentro o hacia fuera de la mesoglea. [22]
Locomoción
La superficie exterior presenta generalmente ocho filas de peines, llamadas placas de natación, que se utilizan para nadar. Las filas están orientadas para correr desde cerca de la boca (el "polo oral") hasta el extremo opuesto (el "polo aboral"), y están espaciadas más o menos uniformemente alrededor del cuerpo, [18] aunque los patrones de espaciado varían según la especie y en la mayoría de las especies las filas de peines se extienden solo una parte de la distancia desde el polo aboral hacia la boca. Los "peines" (también llamados "ctenes" o "placas de peine") corren a lo largo de cada fila, y cada uno consta de miles de cilios inusualmente largos, de hasta 2 milímetros (0,08 pulgadas). A diferencia de los cilios y flagelos convencionales, que tienen una estructura de filamentos dispuestos en un patrón 9 + 2, estos cilios están dispuestos en un patrón 9 + 3, donde se sospecha que el filamento extra compacto tiene una función de soporte. [33] Estos normalmente baten de manera que el golpe de propulsión se aleja de la boca, aunque también pueden invertir la dirección. Por lo tanto, los ctenóforos generalmente nadan en la dirección en la que la boca está comiendo, a diferencia de las medusas . [22] Al intentar escapar de los depredadores, una especie puede acelerar hasta seis veces su velocidad normal; [34] algunas otras especies invierten la dirección como parte de su comportamiento de escape, invirtiendo el movimiento de potencia de los cilios de la placa del peine.
No se sabe con certeza cómo los ctenóforos controlan su flotabilidad, pero los experimentos han demostrado que algunas especies dependen de la presión osmótica para adaptarse al agua de diferentes densidades. [35] Sus fluidos corporales normalmente están tan concentrados como el agua de mar. Si entran en agua salobre menos densa, las rosetas ciliares en la cavidad corporal pueden bombearlos hacia la mesoglea para aumentar su volumen y disminuir su densidad, para evitar hundirse. Por el contrario, si pasan de agua salobre a agua de mar de máxima concentración, las rosetas pueden bombear agua fuera de la mesoglea para reducir su volumen y aumentar su densidad. [22]
Sistema nervioso y sentidos
Los ctenóforos no tienen cerebro ni sistema nervioso central , pero en su lugar tienen una red nerviosa subepidérmica (como una telaraña) que forma un anillo alrededor de la boca y es más densa cerca de estructuras como las hileras de peines, la faringe, los tentáculos (si están presentes) y el complejo sensorial más alejado de la boca. [22] La comunicación entre las células nerviosas utiliza dos métodos diferentes; se ha descubierto que algunas de las neuronas tienen conexiones sinápticas , pero las neuronas de la red nerviosa son muy distintivas al estar fusionadas en un sincitio , en lugar de estar conectadas por sinapsis. Algunos animales fuera de los ctenóforos también tienen células nerviosas fusionadas, pero nunca hasta el punto de formar una red nerviosa completa. [36] [37] [38] Los fósiles muestran que las especies del Cámbrico tenían un sistema nervioso más complejo, con nervios largos que se conectaban con un anillo alrededor de la boca. El único ctenóforo conocido hoy en día con nervios largos es Euplokamis en el orden Cydippida. [39] Sus células nerviosas surgen de las mismas células progenitoras que los coloblastos. [40]
Además, existe una red nerviosa mesogleal menos organizada que consiste en neuritas individuales. La característica sensorial individual más grande es el órgano aboral (en el extremo opuesto de la boca), que está subrayado con su propia red nerviosa. [41] El componente principal de este órgano es un estatocisto , un sensor de equilibrio que consiste en un estatolito, un grano diminuto de carbonato de calcio, sostenido por cuatro haces de cilios , llamados "equilibradores", que detectan su orientación. El estatocisto está protegido por una cúpula transparente hecha de cilios largos e inmóviles. Un ctenóforo no intenta automáticamente mantener el estatolito descansando por igual sobre todos los equilibradores. En cambio, su respuesta está determinada por el "estado de ánimo" del animal, en otras palabras, el estado general del sistema nervioso. Por ejemplo, si un ctenóforo con tentáculos colgantes captura una presa, a menudo pondrá algunas filas de peines en reversa, girando la boca hacia la presa. [22]
Las investigaciones apoyan la hipótesis de que las larvas ciliadas de los cnidarios y los bilaterales comparten un origen antiguo y común. [42] El órgano apical de las larvas está involucrado en la formación del sistema nervioso. [43] El órgano aboral de las medusas peine no es homólogo del órgano apical de otros animales, y por lo tanto la formación de su sistema nervioso tiene un origen embrionario diferente. [44]
Las células nerviosas ctenóforas y el sistema nervioso tienen una bioquímica diferente en comparación con otros animales. Por ejemplo, carecen de los genes y enzimas necesarios para fabricar neurotransmisores como serotonina , dopamina , óxido nítrico , octopamina , noradrenalina y otros, que se observan en todos los demás animales con un sistema nervioso, y faltan los genes que codifican los receptores para cada uno de estos neurotransmisores. [45] La catalasa monofuncional (CAT), una de las tres familias principales de enzimas antioxidantes que se dirigen al peróxido de hidrógeno , una molécula de señalización importante para la actividad sináptica y neuronal, también está ausente, probablemente debido a la pérdida de genes. [46] Se ha descubierto que utilizan L-glutamato como neurotransmisor y tienen una variedad inusualmente alta de receptores de glutamato ionotrópicos y genes para la síntesis y el transporte de glutamato en comparación con otros metazoos. [47] El contenido genómico de los genes del sistema nervioso es el más pequeño conocido de cualquier animal, y podría representar los requisitos genéticos mínimos para un sistema nervioso funcional. [48] El hecho de que partes del sistema nervioso presenten neuronas fusionadas directamente, sin sinapsis, sugiere que los ctenóforos podrían formar un grupo hermano de otros metazoos, habiendo desarrollado un sistema nervioso de forma independiente. [38] Si los ctenóforos son el grupo hermano de todos los demás metazoos, los sistemas nerviosos pueden haberse perdido en esponjas y placozoos, o haber surgido más de una vez entre los metazoos. [49]
Cidípidos
{kind=link}

Los ctenóforos cidípidos tienen cuerpos más o menos redondeados, a veces casi esféricos y otras veces más cilíndricos u ovalados; la grosella espinosa costera común, Pleurobrachia , a veces tiene un cuerpo ovalado con la boca en el extremo estrecho, [22] aunque algunos individuos son más uniformemente redondos. Desde lados opuestos del cuerpo se extiende un par de tentáculos largos y delgados, cada uno alojado en una vaina en la que puede retraerse. [18] Algunas especies de cidípidos tienen cuerpos que están aplanados en diversos grados de modo que son más anchos en el plano de los tentáculos. [22]
Los tentáculos de los ctenóforos cidípidos suelen estar bordeados por tentillas ("pequeños tentáculos"), aunque algunos géneros tienen tentáculos simples sin estas ramas laterales. Los tentáculos y las tentillas están densamente cubiertos de coloblastos microscópicos que capturan a la presa pegándose a ella. Los coloblastos son células especializadas con forma de hongo en la capa externa de la epidermis y tienen tres componentes principales: una cabeza abovedada con vesículas (cámaras) que contienen adhesivo; un tallo que ancla la célula en la capa inferior de la epidermis o en la mesoglea; y un hilo espiral que se enrolla alrededor del tallo y está unido a la cabeza y a la raíz del tallo. La función del hilo espiral es incierta, pero puede absorber la tensión cuando la presa intenta escapar y, por lo tanto, evitar que el coloblasto se desgarre. [22] Una especie, Minictena luteola, que solo mide 1,5 mm de diámetro, tiene cinco tipos diferentes de células coloblastas. [50] [51]
Además de los coloblastos, los miembros del género Haeckelia , que se alimentan principalmente de medusas , incorporan los nematocitos urticantes de sus víctimas en sus propios tentáculos; algunos nudibranquios que se alimentan de cnidarios incorporan de manera similar nematocitos en sus cuerpos para defenderse. [52] Las tentillas de Euplokamis difieren significativamente de las de otros cídipidos: contienen músculo estriado , un tipo de célula desconocido en el filo Ctenophora; y están enrolladas cuando están relajadas, mientras que las tentillas de todos los demás ctenóforos conocidos se alargan cuando están relajadas. Las tentillas de Euplokamis tienen tres tipos de movimiento que se utilizan para capturar presas: pueden salir disparadas muy rápidamente (en 40 a 60 milisegundos ); pueden retorcerse, lo que puede atraer a la presa comportándose como pequeños gusanos planctónicos; y se enrollan alrededor de la presa. El movimiento único es un movimiento de desenrollado impulsado por la contracción del músculo estriado . El movimiento de enrollamiento se produce por músculos lisos , pero de un tipo altamente especializado. El enrollamiento alrededor de la presa se logra en gran medida mediante el retorno de las tentillas a su estado inactivo, pero las espirales pueden apretarse mediante músculos lisos. [53]
Hay ocho filas de peines que van desde cerca de la boca hasta el extremo opuesto, y están espaciadas uniformemente alrededor del cuerpo. [18] Los "peines" baten en un ritmo metacrónico bastante parecido al de una ola mexicana . [54] Desde cada equilibrador en el estatocisto, un surco ciliar sale por debajo de la cúpula y luego se divide para conectarse con dos filas de peines adyacentes, y en algunas especies corre a lo largo de las filas de peines. Esto forma un sistema mecánico para transmitir el ritmo del batido desde los peines a los equilibradores, a través de perturbaciones del agua creadas por los cilios. [55]
Lobatos
{kind=link}
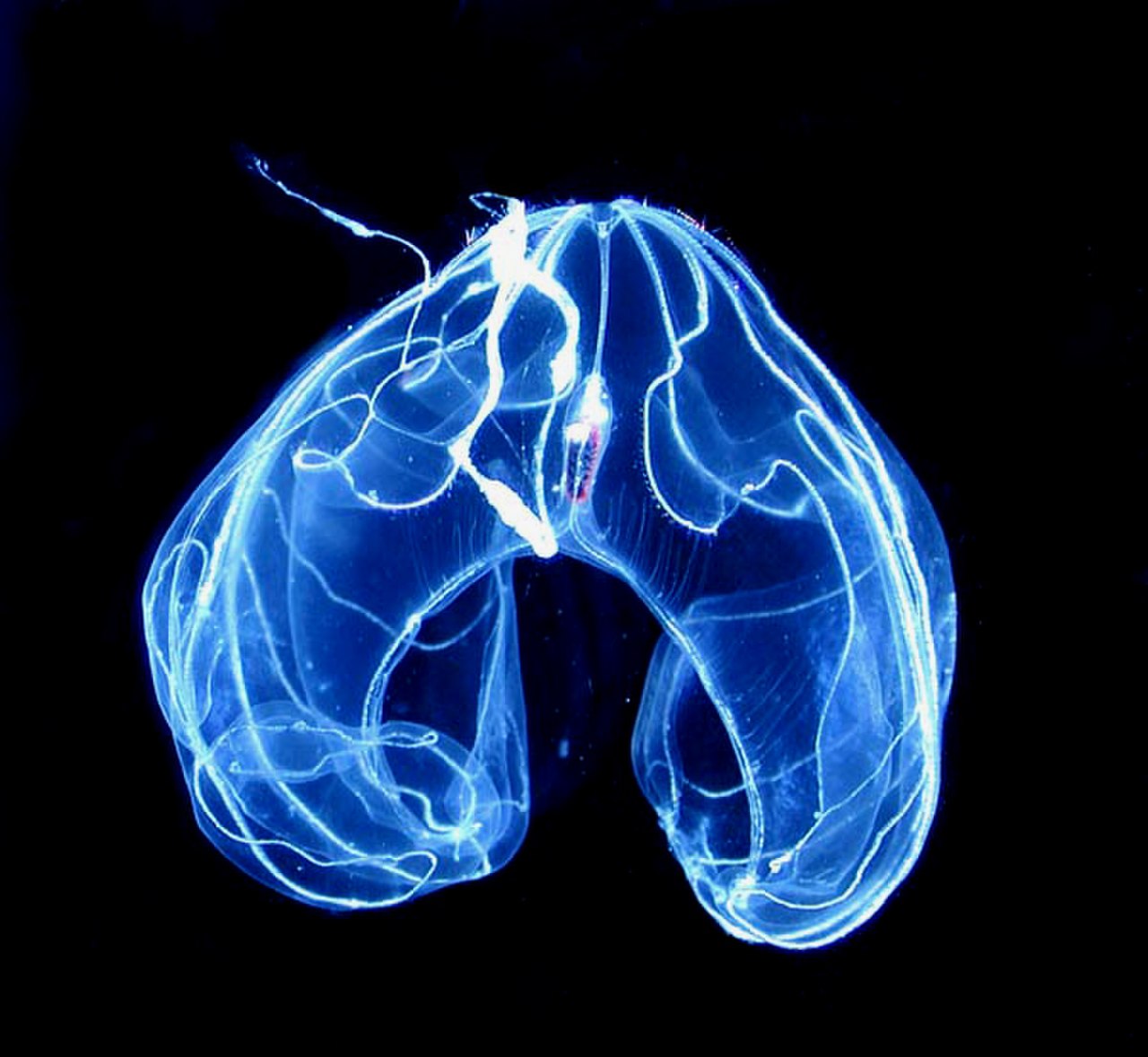
Los lobatos tienen un par de lóbulos, que son extensiones musculares en forma de copa del cuerpo que se proyectan más allá de la boca. Sus discretos tentáculos se originan en las comisuras de la boca, recorriendo surcos enrevesados y extendiéndose sobre la superficie interna de los lóbulos (en lugar de arrastrarse muy atrás, como en los cídipidos). Entre los lóbulos a cada lado de la boca, muchas especies de lobatos tienen cuatro aurículas, proyecciones gelatinosas bordeadas de cilios que producen corrientes de agua que ayudan a dirigir a las presas microscópicas hacia la boca. Esta combinación de estructuras permite a los lobatos alimentarse continuamente de presas planctónicas suspendidas . [22]
Los lobados tienen ocho hileras de peines, que se originan en el polo aboral y por lo general no se extienden más allá del cuerpo hasta los lóbulos; en especies con (cuatro) aurículas, los cilios que bordean las aurículas son extensiones de los cilios en cuatro de las hileras de peines. La mayoría de los lobados son bastante pasivos cuando se mueven a través del agua, utilizando los cilios en sus hileras de peines para propulsarse, [22] aunque Leucothea tiene aurículas largas y activas cuyos movimientos también contribuyen a la propulsión. Los miembros de los géneros lobados Bathocyroe y Ocyropsis pueden escapar del peligro aplaudiendo sus lóbulos, de modo que el chorro de agua expulsado los empuja hacia atrás muy rápidamente. [56] A diferencia de los cídippidos, los movimientos de los peines de los lobados están coordinados por nervios en lugar de por perturbaciones del agua creadas por los cilios, sin embargo, los peines en la misma hilera baten en el mismo estilo de onda mexicana que las hileras de peines coordinadas mecánicamente de los cídippidos y los beroideos. [55] Esto puede haber permitido que los lobados crecieran más que los cidípidos y tuvieran formas menos parecidas a los huevos. [54]
Una especie inusual descrita por primera vez en 2000, Lobatolampea tetragona , ha sido clasificada como lobada, aunque los lóbulos son "primitivos" y el cuerpo es como una medusa cuando flota y como un disco cuando descansa en el fondo del mar. [30]
Beroides
{kind=link}

Los Beroida , también conocidos como Nuda , no tienen apéndices para alimentarse, pero su gran faringe , justo dentro de la gran boca y llenando la mayor parte del cuerpo en forma de saco, tiene "macrocilia" en el extremo oral. Estos haces fusionados de varios miles de cilios grandes son capaces de "morder" trozos de presa que son demasiado grandes para tragarlos enteros, casi siempre otros ctenóforos. [57] Delante del campo de macrocilios, en los "labios" de la boca en algunas especies de Beroe , hay un par de tiras estrechas de células epiteliales adhesivas en la pared del estómago que "cierran" la boca cuando el animal no se está alimentando, formando conexiones intercelulares con la tira adhesiva opuesta. Este cierre hermético agiliza la parte delantera del animal cuando persigue a una presa. [58]
Otras formas corporales
El Ganeshida tiene un par de pequeños lóbulos orales y un par de tentáculos. El cuerpo es circular en sección transversal en lugar de ovalado, y la faringe se extiende sobre las superficies internas de los lóbulos. [22]
Los Thalassocalycida , descubiertos en 1978 y conocidos a partir de una sola especie, [59] son similares a medusas, con cuerpos acortados en la dirección oral-aboral y cortas crestas en la superficie más alejada de la boca, que se originan cerca del polo aboral. Capturan presas mediante movimientos de la campana y posiblemente utilizando dos tentáculos cortos. [22]
Los cestidos ("animales de cinturón") son animales planctónicos con forma de cinta, con la boca y el órgano aboral alineados en el medio de los bordes opuestos de la cinta. Hay un par de hileras de peines a lo largo de cada borde aboral, y una tentilla que emerge de un surco a lo largo del borde oral, que fluye hacia atrás a través de la mayor parte de la superficie corporal similar a un ala. Los cestidos pueden nadar ondulando sus cuerpos, así como mediante el batido de sus hileras de peines. Hay dos especies conocidas, con distribución mundial en aguas cálidas y templadas cálidas: Cestum veneris (" cintura de Venus ") se encuentra entre los ctenóforos más grandes, hasta 1,5 metros (4,9 pies) de largo, y puede ondular lentamente o bastante rápido. Velamen parallelum , que normalmente tiene menos de 20 centímetros (0,66 pies) de largo, puede moverse mucho más rápido en lo que se ha descrito como un "movimiento de dardo". [22] [60]
La mayoría de los platicténidos tienen cuerpos ovalados que se aplanan en la dirección oral-aboral, con un par de tentáculos con tentillas en la superficie aboral. Se adhieren a las superficies y se arrastran por ellas evirtiendo la faringe y utilizándola como un "pie" muscular. Todas las especies de platicténidos conocidas, excepto una, carecen de filas de peines. [22] Los platicténidos suelen tener colores crípticos, viven en rocas, algas o en las superficies corporales de otros invertebrados y a menudo se revelan por sus largos tentáculos con muchas ramas laterales, que se ven fluir desde la parte posterior del ctenóforo hacia la corriente.
Reproducción y desarrollo
{kind=link}

Los adultos de la mayoría de las especies pueden regenerar tejidos dañados o eliminados, [61] aunque solo los platicténidos se reproducen por clonación , separando de los bordes de sus cuerpos planos fragmentos que se convierten en nuevos individuos. [22] Las investigaciones de laboratorio sobre Mnemiopsis leidyi también muestran que cuando a dos individuos se les eliminan partes de sus cuerpos, pueden fusionarse, incluidos sus sistemas nervioso y digestivo, incluso cuando los dos individuos son genéticamente diferentes. Un fenómeno que hasta ahora solo se ha encontrado en medusas peine. [62]
El último ancestro común (LCA) de los ctenóforos era hermafrodita . [63] Algunos son hermafroditas simultáneos, que pueden producir tanto óvulos como espermatozoides al mismo tiempo, mientras que otros son hermafroditas secuenciales, en los que los óvulos y los espermatozoides maduran en diferentes momentos. No hay metamorfosis . [64] Se sabe que al menos tres especies han desarrollado sexos separados ( dioecia ); Ocyropsis crystallina y Ocyropsis maculata en el género Ocyropsis y Bathocyroe fosteri en el género Bathocyroe . [65] Las gónadas se encuentran en las partes de la red de canales internos debajo de las filas de peines, y los óvulos y los espermatozoides se liberan a través de poros en la epidermis. La fertilización es generalmente externa , pero los platicténidos utilizan la fertilización interna y mantienen los huevos en cámaras de cría hasta que eclosionan. Ocasionalmente se ha observado autofecundación en especies del género Mnemiopsis , [22] y se cree que la mayoría de las especies hermafroditas son autofértiles. [19]
El desarrollo de los huevos fertilizados es directo; no hay una forma larvaria distintiva. Los juveniles de todos los grupos son generalmente planctónicos , y la mayoría de las especies se parecen a cídipidos adultos en miniatura, desarrollando gradualmente sus formas corporales adultas a medida que crecen. En el género Beroe , sin embargo, los juveniles tienen bocas grandes y, al igual que los adultos, carecen tanto de tentáculos como de vainas de tentáculos. En algunos grupos, como los platicténidos planos que viven en el fondo, los juveniles se comportan más como larvas verdaderas. Viven entre el plancton y, por lo tanto, ocupan un nicho ecológico diferente al de sus padres, y solo alcanzan la forma adulta mediante una ontogenia más radical [22] después de caer al fondo marino. [19]
Al menos en algunas especies, los ctenóforos juveniles parecen capaces de producir pequeñas cantidades de óvulos y espermatozoides mientras están muy por debajo del tamaño adulto, y los adultos producen óvulos y espermatozoides mientras tengan suficiente alimento. Si se quedan sin alimento, primero dejan de producir óvulos y espermatozoides, y luego disminuyen de tamaño. Cuando el suministro de alimento mejora, vuelven a crecer hasta su tamaño normal y luego reanudan la reproducción. Estas características hacen que los ctenóforos sean capaces de aumentar sus poblaciones muy rápidamente. [19] Los miembros de Lobata y Cydippida también tienen una forma de reproducción llamada disogenia; dos etapas de madurez sexual, primero como larva y luego como juveniles y adultos. Durante su tiempo como larvas son capaces de liberar gametos periódicamente. Una vez que termina su primer período reproductivo, no volverán a producir más gametos hasta más tarde. Una población de Mertensia ovum en el mar Báltico central se ha vuelto pedogenética y consiste únicamente en larvas sexualmente maduras de menos de 1,6 mm. [66] [67]
En Mnemiopsis leidyi , la señalización del óxido nítrico (NO) está presente tanto en los tejidos adultos como en las etapas embrionarias posteriores, lo que sugiere la participación del NO en los mecanismos de desarrollo. [68] La forma madura de la misma especie también puede volver a la etapa de cidípido cuando se ve afectada por factores estresantes ambientales. [69]
Colores y bioluminiscencia
{kind=link}

La mayoría de los ctenóforos que viven cerca de la superficie son en su mayoría incoloros y casi transparentes. Sin embargo, algunas especies que viven a mayor profundidad están fuertemente pigmentadas, por ejemplo, la especie conocida como "Tortugas rojas" [70] (ver ilustración aquí), que aún no ha sido descrita formalmente. [19] Los platicténidos generalmente viven adheridos a otros organismos del fondo marino y a menudo tienen colores similares a estos organismos hospedadores. [19] El intestino del género de aguas profundas Bathocyroe es rojo, lo que oculta la bioluminiscencia de los copépodos que ha tragado. [56]
Las filas de peines de la mayoría de los ctenóforos planctónicos producen un efecto arco iris, que no es causado por la bioluminiscencia sino por la dispersión de la luz a medida que los peines se mueven. [19] [71] La mayoría de las especies también son bioluminiscentes, pero la luz suele ser azul o verde y solo se puede ver en la oscuridad. [19] Sin embargo, algunos grupos importantes, incluidos todos los platicténidos conocidos y el género de cídipidos Pleurobrachia , son incapaces de bioluminiscencia. [72]
Cuando algunas especies, entre ellas Bathyctena chuni , Euplokamis stationis y Eurhamphaea vexilligera , son molestadas, producen secreciones (tinta) que emiten luminiscencia en longitudes de onda muy similares a las de sus cuerpos. Los ejemplares jóvenes emiten una luminiscencia más intensa en relación con el tamaño de su cuerpo que los adultos, cuya luminiscencia se difunde por todo el cuerpo. Una investigación estadística detallada no ha sugerido la función de la bioluminiscencia de los ctenóforos ni ha producido ninguna correlación entre su color exacto y ningún aspecto del entorno de los animales, como la profundidad o si viven en aguas costeras o en medio del océano. [73]
En los ctenóforos, la bioluminiscencia es causada por la activación de proteínas activadas por calcio llamadas fotoproteínas en células llamadas fotocitos , que a menudo están confinadas a los canales meridionales que se encuentran debajo de las ocho filas de peines. En el genoma de Mnemiopsis leidyi, diez genes codifican fotoproteínas. Estos genes se coexpresan con genes de opsina en los fotocitos en desarrollo de Mnemiopsis leidyi , lo que plantea la posibilidad de que la producción de luz y la detección de luz puedan estar trabajando juntas en estos animales. [74]
Ecología
{kind=link}

Distribución
Los ctenóforos se encuentran en la mayoría de los ambientes marinos: desde aguas polares a -2 °C hasta los trópicos a 30 °C; cerca de las costas y en medio del océano; desde las aguas superficiales hasta las profundidades del océano a más de 7000 metros. [75] Los mejor conocidos son los géneros Pleurobrachia , Beroe y Mnemiopsis , ya que estas formas costeras planctónicas se encuentran entre las que es más probable que se recolecten cerca de la costa. [31] [56] No se han encontrado ctenóforos en agua dulce.
En 2013, se registró Mnemiopsis en el lago Birket Qarun y en 2014 en el lago El Rayan II, ambos cerca de Faiyum en Egipto, donde se introdujeron accidentalmente por el transporte de alevines de peces (salmonete). Aunque muchas especies prefieren aguas salobres como estuarios y lagunas costeras en conexión abierta con el mar, este fue el primer registro de un entorno interior. Ambos lagos son salinos, siendo Birket Qarun hipersalino, y muestra que algunos ctenóforos pueden establecerse en entornos límnicos salinos sin conexión con el océano. A largo plazo, no se espera que las poblaciones sobrevivan. Los dos factores limitantes en los lagos salinos son la disponibilidad de alimentos y una dieta variada, y las altas temperaturas durante los veranos calurosos. Debido a que un isópodo parásito, Livoneca redmanii , se introdujo al mismo tiempo, es difícil decir qué parte del impacto ecológico de las especies invasoras es causado solo por el ctenóforo. [76] [77]
Los ctenóforos pueden ser abundantes durante los meses de verano en algunas localidades costeras, pero en otros lugares son poco comunes y difíciles de encontrar.
En las bahías donde se encuentran en cantidades muy elevadas, la depredación por parte de los ctenóforos puede controlar las poblaciones de pequeños organismos zooplanctónicos, como los copépodos , que de otro modo podrían acabar con el fitoplancton (plantas planctónicas), que son una parte vital de las cadenas alimentarias marinas .
Presas y depredadores
Casi todos los ctenóforos son depredadores : no hay vegetarianos y solo un género que es parcialmente parásito . [56] Si la comida es abundante, pueden comer 10 veces su propio peso por día. [78] Mientras que Beroe se alimenta principalmente de otros ctenóforos, otras especies de aguas superficiales se alimentan de zooplancton (animales planctónicos) que varían en tamaño desde los microscópicos, incluidas las larvas de moluscos y peces, hasta pequeños crustáceos adultos como copépodos , anfípodos e incluso krill . Los miembros del género Haeckelia se alimentan de medusas e incorporan los nematocistos (células urticantes) de sus presas en sus propios tentáculos en lugar de coloblastos . [19] Los ctenóforos han sido comparados con las arañas en su amplia gama de técnicas para capturar presas: algunos cuelgan inmóviles en el agua usando sus tentáculos como "telarañas", algunos son depredadores de emboscada como las arañas saltadoras Salticid , y algunos cuelgan una gota pegajosa en el extremo de un hilo fino, como lo hacen las arañas bolas . Esta variedad explica la amplia gama de formas corporales en un filo con bastante pocas especies. [56] El "cidípido" Lampea de dos tentáculos se alimenta exclusivamente de salpas , parientes cercanos de las ascidias que forman grandes colonias flotantes en forma de cadena, y los juveniles de Lampea se adhieren como parásitos a las salpas que son demasiado grandes para que las traguen. [56] Los miembros del género cydippid Pleurobrachia y el lobado Bolinopsis a menudo alcanzan altas densidades de población en el mismo lugar y tiempo porque se especializan en diferentes tipos de presas: los largos tentáculos de Pleurobrachia capturan principalmente nadadores relativamente fuertes como copépodos adultos, mientras que Bolinopsis generalmente se alimenta de nadadores más pequeños y débiles como rotíferos y larvas de moluscos y crustáceos . [79]
Los ctenóforos solían considerarse "callejones sin salida" en las cadenas alimentarias marinas porque se pensaba que su baja proporción de materia orgánica con respecto a la sal y el agua los convertía en una dieta deficiente para otros animales. También suele ser difícil identificar los restos de ctenóforos en los intestinos de posibles depredadores, aunque a veces las crestas permanecen intactas el tiempo suficiente para proporcionar una pista. Una investigación detallada del salmón chum , Oncorhynchus keta , mostró que estos peces digieren los ctenóforos 20 veces más rápido que un peso equivalente de camarones , y que los ctenóforos pueden proporcionar una buena dieta si hay suficientes de ellos alrededor. Los beroides se alimentan principalmente de otros ctenóforos. Algunas medusas y tortugas comen grandes cantidades de ctenóforos, y las medusas pueden acabar temporalmente con las poblaciones de ctenóforos. Dado que los ctenóforos y las medusas a menudo tienen grandes variaciones estacionales en la población, la mayoría de los peces que se alimentan de ellos son generalistas y pueden tener un mayor efecto en las poblaciones que los comedores de medusas especialistas. Esto se ve subrayado por una observación de peces herbívoros que se alimentan deliberadamente de zooplancton gelatinoso durante las floraciones en el Mar Rojo. [80] Las larvas de algunas anémonas de mar son parásitos de los ctenóforos, al igual que las larvas de algunos platelmintos que parasitan a los peces cuando alcanzan la edad adulta. [81]
Impactos ecológicos
La mayoría de las especies son hermafroditas y los ejemplares jóvenes de al menos algunas especies son capaces de reproducirse antes de alcanzar el tamaño y la forma de los adultos. Esta combinación de hermafroditismo y reproducción temprana permite que las poblaciones pequeñas crezcan a un ritmo explosivo.
{kind=link}

Los ctenóforos pueden equilibrar los ecosistemas marinos al evitar que una sobreabundancia de copépodos se coma todo el fitoplancton (plantas planctónicas), [82] que son los principales productores marinos de materia orgánica a partir de ingredientes no orgánicos. [83]
Por otra parte, a finales de los años 1980 el ctenóforo del Atlántico occidental Mnemiopsis leidyi fue introducido accidentalmente en el Mar Negro y el Mar de Azov a través de los tanques de lastre de los barcos, y ha sido culpado de causar fuertes caídas en las capturas de peces al comer tanto larvas de peces como pequeños crustáceos que de otro modo alimentarían a los peces adultos. [82] Mnemiopsis está bien equipado para invadir nuevos territorios (aunque esto no se predijo hasta después de que colonizara con tanto éxito el Mar Negro), ya que puede reproducirse muy rápidamente y tolerar una amplia gama de temperaturas y salinidades del agua . [84] El impacto se incrementó por la sobrepesca crónica y por la eutrofización que dio a todo el ecosistema un impulso a corto plazo, haciendo que la población de Mnemiopsis aumentara incluso más rápido de lo normal [85] - y sobre todo por la ausencia de depredadores eficientes en estos ctenóforos introducidos. [84] Las poblaciones de Mnemiopsis en esas áreas finalmente fueron controladas por la introducción accidental del ctenóforo norteamericano Beroe ovata , que se alimenta de Mnemiopsis , [86] y por un enfriamiento del clima local entre 1991 y 1993, [85] que desaceleró significativamente el metabolismo del animal. [84] Sin embargo, parece poco probable que la abundancia de plancton en el área se restablezca a los niveles previos a Mnemiopsis . [87]
A finales de los años 1990, Mnemiopsis apareció en el mar Caspio . Beroe ovata llegó poco después y se espera que reduzca, pero no elimine, el impacto de Mnemiopsis allí. Mnemiopsis también llegó al Mediterráneo oriental a finales de los años 1990 y ahora parece estar prosperando en el mar del Norte y el mar Báltico . [19]
Taxonomía
El número de especies de ctenóforos vivos conocidas es incierto, ya que muchas de las que recibieron nombre y se describieron formalmente resultaron ser idénticas a especies conocidas con otros nombres científicos. Claudia Mills estima que existen alrededor de 100 a 150 especies válidas que no son duplicadas, y que al menos otras 25, en su mayoría formas de aguas profundas, han sido reconocidas como distintas, pero aún no se han analizado con suficiente detalle como para respaldar una descripción y denominación formales. [70]
Clasificación temprana
Los primeros escritores combinaron los ctenóforos con los cnidarios en un solo filo llamado Coelenterata debido a las similitudes morfológicas entre los dos grupos. Al igual que los cnidarios, los cuerpos de los ctenóforos consisten en una masa de gelatina, con una capa de células en el exterior y otra que recubre la cavidad interna. En los ctenóforos, sin embargo, estas capas tienen dos células de profundidad, mientras que las de los cnidarios tienen solo una célula de profundidad. Los ctenóforos también se parecen a los cnidarios en que dependen del flujo de agua a través de la cavidad corporal tanto para la digestión como para la respiración, así como en tener una red nerviosa descentralizada en lugar de un cerebro. Los estudios genómicos han sugerido que las neuronas de Ctenophora, que difieren en muchos aspectos de las neuronas de otros animales, evolucionaron independientemente de las de los otros animales, [88] y la creciente conciencia de las diferencias entre las medusas peine y los otros celentáridos ha persuadido a los autores más recientes a clasificar los dos como filos separados . La posición de los ctenóforos en el árbol genealógico evolutivo de los animales ha sido debatida durante mucho tiempo, y la opinión mayoritaria en la actualidad, basada en la filogenética molecular , es que los cnidarios y los bilaterales están más estrechamente relacionados entre sí que con los ctenóforos.
Taxonomía moderna
{kind=link}

La clasificación tradicional divide a los ctenóforos en dos clases , los que tienen tentáculos ( Tentaculata ) y los que no los tienen ( Nuda ). La Nuda contiene sólo un orden ( Beroida ) y una familia ( Beroidae ), y dos géneros , Beroe (varias especies) y Neis (una especie). [70]
Los Tentaculata se dividen en los ocho órdenes siguientes : [70]
- Cydippida , animales con forma de huevo y tentáculos largos [22]
- Lobata , con lóbulos gruesos pareados [22]
- Platyctenida , animales aplanados que viven en el fondo del mar o cerca de él; la mayoría carece de crestas cuando son adultos y utilizan sus faringes como ventosas para adherirse a las superficies [22]
- Ganeshida , con un par de pequeños lóbulos alrededor de la boca, pero una faringe extendida como la de los platicténidos [22]
- Cambojiida
- Criptolobiferidos
- Thalassocalycida , con tentáculos cortos y un "paraguas" parecido al de una medusa [22]
- Cestida , con forma de cinta y los ctenóforos más grandes [22]
Existen géneros fósiles que se consideran grupo troncal .
Historia evolutiva
A pesar de sus cuerpos frágiles y gelatinosos, se han encontrado fósiles en Lagerstätten que se cree que representan ctenóforos (aparentemente sin tentáculos pero con muchas más hileras de peines que las formas modernas) ya en el Cámbrico temprano , hace unos 515 millones de años . Sin embargo, un análisis filogenético molecular reciente concluye que el ancestro común se originó hace aproximadamente 350 millones de años ± 88 millones de años, lo que contradice las estimaciones anteriores que sugieren que ocurrió hace 66 millones de años después del evento de extinción del Cretácico-Paleógeno . [89]
Registro fósil
Debido a sus cuerpos blandos y gelatinosos, los ctenóforos son extremadamente raros como fósiles, y los fósiles que se han interpretado como ctenóforos solo se han encontrado en lagerstätten , lugares donde el ambiente era excepcionalmente adecuado para la preservación de tejido blando. Hasta mediados de la década de 1990, solo se conocían dos especímenes lo suficientemente buenos para el análisis, ambos miembros del grupo de la corona, del período Devónico temprano (Emsiano) . Luego se encontraron tres especies putativas adicionales en Burgess Shale y otras rocas canadienses de edad similar, hace unos 505 millones de años en el período Cámbrico medio . Los tres carecían de tentáculos, pero tenían entre 24 y 80 filas de peines, mucho más que las ocho típicas de las especies actuales. También parecen haber tenido estructuras internas similares a órganos diferentes a todo lo encontrado en ctenóforos vivos. Una de las especies fósiles reportadas por primera vez en 1996 tenía una boca grande, aparentemente rodeada por un borde doblado que puede haber sido muscular. [3] La evidencia de China un año después sugiere que tales ctenóforos estaban muy extendidos en el Cámbrico, pero tal vez eran muy diferentes de las especies modernas; por ejemplo, las filas de peines de un fósil estaban montadas sobre veletas prominentes. [90] El fósil más joven de una especie fuera del grupo corona es la especie Daihuoides del Devónico tardío, que pertenece a un grupo basal que se suponía que se había extinguido más de 140 millones de años antes. [91]
El Eoandromeda ediacárico podría representar putativamente una medusa peine. [4] Tiene simetría óctuple, con ocho brazos espirales que se asemejan a las filas en forma de peine de un ctenóforo. Si es de hecho un ctenóforo, coloca al grupo cerca del origen de Bilateria. [92] El fósil sésil con forma de fronda del Cámbrico temprano Stromatoveris , del lagerstätte de Chengjiang en China y que data de hace unos 515 millones de años , es muy similar a Vendobionta del período Ediacárico precedente. De-Gan Shu, Simon Conway Morris , et al. encontraron en sus ramas lo que consideraron filas de cilios, utilizados para la alimentación por filtración . Sugirieron que Stromatoveris era una "tía" evolutiva de los ctenóforos, y que los ctenóforos se originaron a partir de animales sésiles cuyos descendientes se convirtieron en nadadores y cambiaron los cilios de un mecanismo de alimentación a un sistema de propulsión. [93] Otros fósiles cámbricos que apoyan la idea de que los ctenóforos evolucionaron a partir de formas sésiles son Dinomischus , Daihua , Xianguangia y Siphusauctum , que también vivieron en el fondo marino, tenían esqueletos orgánicos y tentáculos cubiertos de cilios que rodeaban su boca, que se han encontrado mediante análisis cladístico como miembros del grupo madre de los ctenóforos [94] [95]
Los fósiles cámbricos de 520 millones de años también de Chengjiang en China muestran una clase ahora completamente extinta de ctenóforo, llamada " Scleroctenophora ", que tenía un esqueleto interno complejo con espinas largas. [96] El esqueleto también sostenía ocho aletas de cuerpo blando, que podrían haber sido utilizadas para nadar y posiblemente alimentarse. Una forma, Thaumactena , tenía un cuerpo aerodinámico parecido al de los gusanos flecha y podría haber sido un nadador ágil. [5]
Relación con otros grupos de animales
La relación filogenética de los ctenóforos con el resto de metazoos es muy importante para nuestra comprensión de la evolución temprana de los animales y el origen de la multicelularidad. Ha sido el foco de debate durante muchos años. Se ha propuesto que los ctenóforos sean el linaje hermano de los Bilateria , [97] [98] hermano de los Cnidaria , [99] [100] [101] [102] hermano de Cnidaria , Placozoa y Bilateria , [103] [104] [105] y hermano de todos los demás animales. [9] [106]
Walter Garstang en su libro Formas larvarias y otros versos zoológicos ( Mülleria y el ctenóforo ) incluso expresó una teoría de que los ctenóforos descendían de una larva neoténica de Mülleria de un policlado .
Una serie de estudios que analizaron la presencia y ausencia de miembros de familias de genes y vías de señalización (por ejemplo, homeoboxes , receptores nucleares , la vía de señalización Wnt y canales de sodio ) mostraron evidencia congruente con los dos últimos escenarios, de que los ctenóforos son hermanos de Cnidaria , Placozoa y Bilateria o hermanos de todos los demás filos animales. [107] [108] [109] [110] Varios estudios más recientes que comparan genomas secuenciados completos de ctenóforos con otros genomas animales secuenciados también han apoyado a los ctenóforos como el linaje hermano de todos los demás animales. [111] [28] [112] [113] Esta posición sugeriría que los tipos de células neuronales y musculares se perdieron en los principales linajes animales (por ejemplo, Porifera y Placozoa ) o evolucionaron independientemente en el linaje de los ctenóforos. [111]
Otros investigadores han argumentado que la colocación de Ctenophora como hermana de todos los demás animales es una anomalía estadística causada por la alta tasa de evolución de los genomas de los ctenóforos, y que Porifera (esponjas) es en cambio el taxón animal que diverge más temprano. [105] [114] [115] [116] [117] También tienen tasas extremadamente altas de evolución mitocondrial , [118] y el contenido de ARN/proteína más pequeño conocido del genoma de ADNmt en animales. [119] Como tal, los Ctenophora parecen ser un clado diploblástico basal . De acuerdo con este último punto, el análisis de una alineación de secuencias muy grande a escala taxonómica de los metazoos (1719 proteínas que totalizan c. 400 000 posiciones de aminoácidos ) mostró que los ctenóforos emergen como el segundo linaje animal de ramificación más temprana, y las esponjas son un grupo hermano de todos los demás animales multicelulares. [8] Además, la investigación sobre los genes de la mucina , que permiten a un animal producir moco, muestra que las esponjas nunca los han tenido mientras que todos los demás animales, incluidas las medusas peine, parecen compartir genes con un origen común. [120] Y se ha revelado que a pesar de todas sus diferencias, las neuronas de los ctenóforos comparten la misma base que las neuronas de los cnidarios después de que los hallazgos mostraran que las neuronas que expresan péptidos son probablemente ancestrales de los neurotransmisores químicos. [121]
Otro estudio rechaza firmemente la hipótesis de que las esponjas son el grupo hermano de todos los demás animales existentes y establece la ubicación de Ctenophora como el grupo hermano de todos los demás animales, y el desacuerdo con el último artículo mencionado se explica por problemas metodológicos en los análisis de ese trabajo. [13] Ni los ctenóforos ni las esponjas poseen vías HIF , [122] su genoma expresa solo un único tipo de canal de calcio dependiente de voltaje a diferencia de otros animales que tienen tres tipos, [123] y son los únicos filos animales conocidos que carecen de genes hox verdaderos . [28] Unas pocas especies de otros filos; la larva del pilidium nemertino , la larva de la especie Phoronid Phoronopsis harmeri y la larva del gusano bellota Schizocardium californicum , tampoco dependen de los genes hox en su desarrollo larvario, pero los necesitan durante la metamorfosis para alcanzar su forma adulta. [124] [125] [126] Los genes de innexina , que codifican proteínas utilizadas para la comunicación intercelular en animales, también parecen haber evolucionado independientemente en los ctenóforos. [127]
Relaciones dentro de Ctenophora
| ||||||||||||||||||||||||||||
Dado que todos los ctenóforos modernos, excepto los beroideos, tienen larvas similares a los cidípidos, se ha asumido ampliamente que su último ancestro común también se parecía a los cidípidos, ya que tenía un cuerpo con forma de huevo y un par de tentáculos retráctiles. El análisis puramente morfológico de Richard Harbison en 1985 concluyó que los cidípidos no son monofiléticos, es decir, no contienen todos y solo los descendientes de un único ancestro común que era en sí un cidípido. En cambio, descubrió que varias familias de cidípidos eran más similares a los miembros de otros órdenes de ctenóforos que a otros cidípidos. También sugirió que el último ancestro común de los ctenóforos modernos era similar a un cidípido o a un beroideo. [129] Un análisis de filogenia molecular en 2001, usando 26 especies, incluyendo 4 recientemente descubiertas, confirmó que los cídipidos no son monofiléticos y concluyó que el último ancestro común de los ctenóforos modernos era similar a los cídipidos. También encontró que las diferencias genéticas entre estas especies eran muy pequeñas, tan pequeñas que las relaciones entre Lobata, Cestida y Thalassocalycida seguían siendo inciertas. Esto sugiere que el último ancestro común de los ctenóforos modernos fue relativamente reciente, y tal vez sobrevivió al evento de extinción del Cretácico-Paleógeno hace 65,5 millones de años , mientras que otros linajes perecieron. Cuando el análisis se amplió para incluir representantes de otros filos , concluyó que los cnidarios probablemente están más estrechamente relacionados con los bilaterales que cualquiera de los grupos con los ctenóforos, pero que este diagnóstico es incierto. [128] Un clado que incluye a Mertensia , Charistephane y Euplokamis puede ser el linaje hermano de todos los demás ctenóforos. [130] [13]
Los tiempos de divergencia estimados a partir de datos moleculares indicaron aproximadamente hace cuántos millones de años (Mya) se diversificaron los principales clados: 350 Mya para Cydippida en relación con otros Ctenophora, y 260 Mya para Platyctenida en relación con Beroida y Lobata. [13]
Véase también
Referencias
- ^ Chen, Jun-Yuan; Schopf, J. William; Bottjer, David J.; Zhang, Chen-Yu; Kudryavtsev, Anatoliy B.; Tripathi, Abhishek B.; et al. (abril de 2007). "Espectros Raman de un embrión de ctenóforo del Cámbrico Inferior del suroeste de Shaanxi, China". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 104 (15): 6289–6292. Bibcode :2007PNAS..104.6289C. doi : 10.1073/pnas.0701246104 . PMC 1847456 . PMID 17404242.
- ^ Stanley, GD; Stürmer, W. (9 de junio de 1983). "El primer ctenóforo fósil del Devónico inferior de Alemania Occidental". Nature . 303 (5917): 518–520. Bibcode :1983Natur.303..518S. doi :10.1038/303518a0. S2CID 4259485.
- ^ ab Conway Morris, S. ; Collins, DH (29 de marzo de 1996). "Ctenóforos del Cámbrico Medio de la Formación Stephen, Columbia Británica, Canadá". Philosophical Transactions of the Royal Society B: Biological Sciences . 351 (1337): 279–308. Bibcode :1996RSPTB.351..279C. doi :10.1098/rstb.1996.0024.
- ^ ab Tang, F.; Bengtson, S.; Wang, Y.; Wang, XL; Yin, CY (20 de septiembre de 2011). "Eoandromeda y el origen de Ctenophora". Evolución y desarrollo . 13 (5): 408–414. doi :10.1111/j.1525-142X.2011.00499.x. PMID 23016902. S2CID 28369431.
- ^ ab Shu, Degan; Zhang, Zhifei; Zhang, colmillo; Sol, Ge; Han, Jian; Xiao, Shuhai; Ou, Qiang (julio de 2015). "Una historia desaparecida de esqueletización en medusas de peine del Cámbrico". Avances científicos . 1 (6): e1500092. Código Bib : 2015SciA....1E0092O. doi :10.1126/sciadv.1500092. PMC 4646772 . PMID 26601209.
- ^ Fowler, George Herbert (1911). . En Chisholm, Hugh (ed.). Encyclopædia Britannica . Vol. 7 (11.ª ed.). Cambridge University Press. pág. 593.
- ^ Registro Mundial de Especies Marinas (base de datos). VLIZ. 18 de septiembre de 2019. Consultado el 19 de febrero de 2024 .
- ^ ab Simion, Paul; Philippe, Hervé; Baurain, Denis; Jager, Muriel; Richter, Daniel J.; diFranco, Arnaud; et al. (2017). "Un conjunto de datos filogenómicos amplio y consistente respalda a las esponjas como el grupo hermano de todos los demás animales". Current Biology . 27 (7): 958–967. Bibcode :2017CBio...27..958S. doi : 10.1016/j.cub.2017.02.031 . PMID 28318975.
- ^ ab Dunn, Casey W.; Hejnol, Andreas; Matus, David Q.; Pang, Kevin; Browne, William E.; Smith, Stephen A.; et al. (2008). "El muestreo filogenómico amplio mejora la resolución del árbol de la vida animal". Nature . 452 (7188): 745–749. Bibcode :2008Natur.452..745D. doi :10.1038/nature06614. PMID 18322464. S2CID 4397099.
- ^ Baxevanis, Andreas D.; Martindale, Mark Q.; Mullikin, James C.; Wolfsberg, Tyra G.; Dunn, Casey W.; Haddock, Steven HD; et al. (13 de diciembre de 2013). "El genoma del ctenóforo Mnemiopsis leidyi y sus implicaciones para la evolución del tipo celular". Science . 342 (6164): 1242592. doi :10.1126/science.1242592. PMC 3920664 . PMID 24337300.
- ^ Pisani, Davide; Pett, Walker; Dohrmann, Martin; Feuda, Roberto; Rota-Stabelli, Omar; Philippe, Hervé; et al. (2015). "Los datos genómicos no respaldan que las medusas peine sean el grupo hermano de todos los demás animales". Actas de la Academia Nacional de Ciencias de los Estados Unidos . 112 (50): 15402–15407. Bibcode :2015PNAS..11215402P. doi : 10.1073/pnas.1518127112 . PMC 4687580 . PMID 26621703.
- ^ Berwald, Juli (noviembre de 2017). Spineless: The science of jellyfish and the art of growing a backbone (Sin espinas: la ciencia de las medusas y el arte de desarrollar una columna vertebral ). Ivanyi, Rachel (ilus.) (1.ª edición en formato de libro de alta definición). Nueva York, NY: Riverhead Books (Penguin Random House). ISBN 978-0735211261. Número de serie LCCN 2017005838.
- ^ abcd Whelan, Nathan V.; Kocot, Kevin M.; Moroz, Tatiana P.; Mukherjee, Krishanu; Williams, Peter; Paulay, Gustav; et al. (noviembre de 2017). "Relaciones de los ctenóforos y su ubicación como grupo hermano de todos los demás animales". Nature Ecology & Evolution . 1 (11): 1737–1746. Bibcode :2017NatEE...1.1737W. doi :10.1038/s41559-017-0331-3. PMC 5664179 . PMID 28993654.
- ^ Halanych, Kenneth M.; Whelan, Nathan V.; Kocot, Kevin M.; Kohn, Andrea B.; Moroz, Leonid L. (9 de febrero de 2016). "Los errores extravían las esponjas". Actas de la Academia Nacional de Ciencias de los Estados Unidos . 113 (8): E946–E947. Bibcode :2016PNAS..113E.946H. doi : 10.1073/pnas.1525332113 . ISSN 0027-8424. PMC 4776479 . PMID 26862177.
- ^ Telford, Maximilian J.; Moroz, Leonid L.; Halanych, Kenneth M. (enero de 2016). "Una disputa entre hermanas". Nature . 529 (7586): 286–287. doi : 10.1038/529286a . ISSN 1476-4687. PMID 26791714. S2CID 4447056.
- ^ Schultz, Darrin T.; Haddock, Steven HD; Bredeson, Jessen V.; Green, Richard E.; Simakov, Oleg; Rokhsar, Daniel S. (17 de mayo de 2023). "Los vínculos genéticos antiguos respaldan a los ctenóforos como hermanos de otros animales". Nature . 618 (7963): 110–117. Bibcode :2023Natur.618..110S. doi :10.1038/s41586-023-05936-6. ISSN 1476-4687. PMC 10232365 . PMID 37198475.
- ^ Ryan, Joseph F.; Schnitzler, Christine E.; Tamm, Sidney L. (diciembre de 2016). "Informe de la reunión de Ctenopalooza: la primera reunión internacional de ctenofórólogos". EvoDevo . 7 (1): 19. doi : 10.1186/s13227-016-0057-3 . hdl : 1912/8430 . S2CID 931968.
- ^ abcdefghijkl Hinde, RT (1998). "Los cnidarios y los ctenóforos". En Anderson, DT (ed.). Zoología de invertebrados . Oxford University Press. págs. 28–57. ISBN
978-0-19-551368-4. - ^ abcdefghijklmn Mills, CE "Ctenóforos: algunas notas de un experto". Seattle, WA: Universidad de Washington . Consultado el 5 de febrero de 2009 .
- ^ de Ruppert, EE; Fox, RS y Barnes, RD (2004). Zoología de invertebrados (7.ª ed.). Brooks / Cole. págs. 111–124. ISBN 978-0-03-025982-1.
- ^ ab Seipel, K.; Schmid, V. (junio de 2005). "Evolución del músculo estriado: medusas y el origen de la triploblastia". Biología del desarrollo . 282 (1): 14–26. doi : 10.1016/j.ydbio.2005.03.032 . PMID 15936326.
- ^ abcdefghijklmnopqrstu vwxyz aa ab ac ad ae af ag ah ai aj Ruppert, EE; Fox, RS y Barnes, RD (2004). Zoología de invertebrados (7.ª ed.). Brooks / Cole. págs. 182–195. ISBN
978-0-03-025982-1. - ^ Moroz, Leonid L.; Norekian, Tigran P. (16 de agosto de 2018). "Atlas de Organización Neuromuscular en el Ctenóforo, Pleurobrachia bachei (A. Agassiz, 1860) ". bioRxiv 10.1101/385435 .
- ^ Viitasalo, S.; Lehtiniemi, M. y Katajisto, T. (2008). "El ctenóforo invasor Mnemiopsis leidyi pasa el invierno en grandes cantidades en el mar Báltico subártico". Journal of Plankton Research . 30 (12): 1431–1436. doi :10.1093/plankt/fbn088.
- ^ Trumble, W.; Brown, L. (2002). "Ctenóforo". Diccionario Oxford de inglés más breve . Oxford University Press.
- ^ Ruppert, EE; Fox, RS y Barnes, RD (2004). Zoología de invertebrados (7.ª ed.). Brooks / Cole. pp. 76–97. ISBN
978-0-03-025982-1. - ^ Bergquist, PR (1998). "Porifera". En Anderson, DT (ed.). Zoología de invertebrados . Oxford University Press. págs. 10-27. ISBN 978-0-19-551368-4.
- ^ abc Moroz, Leonid L.; Kocot, Kevin M.; Citarella, Mathew R.; Dosung, Sohn; Norekian, Tigran P.; Povolotskaya, Inna S.; et al. (junio de 2014). "El genoma del ctenóforo y los orígenes evolutivos de los sistemas neuronales". Nature . 510 (7503): 109–114. Bibcode :2014Natur.510..109M. doi :10.1038/nature13400. PMC 4337882 . PMID 24847885.
- ^ Exposito, JY.; Cluzel, C.; Garrone, R. y Lethias, C. (2002). "Evolución de los colágenos". The Anatomical Record Part A: Discoveries in Molecular, Cellular, and Evolutionary Biology . 268 (3): 302–316. doi : 10.1002/ar.10162 . PMID 12382326. S2CID 12376172.
- ^ ab Horita, T. (marzo de 2000). "Un ctenóforo lobulado no descrito, Lobatolampea tetragona gen. nov. y spec. nov., que representa una nueva familia, de Japón". Zoológico de Mededelingen . 73 (30): 457–464 . Consultado el 3 de enero de 2009 .
- ^ ab Haddock, Steven HD (noviembre de 2004). "Una edad de oro de la gelata: investigación pasada y futura sobre ctenóforos planctónicos y cnidarios". Hydrobiologia . 530–531 (1–3): 549–556. doi :10.1007/s10750-004-2653-9. S2CID 17105070.
- ^ Martindale, MQ; Henry, JQ (octubre de 1999). "El mapeo del destino intracelular en un metazoo basal, el ctenóforo Mnemiopsis leidyi, revela los orígenes del mesodermo y la existencia de linajes celulares indeterminados". Biología del desarrollo . 214 (2): 243–257. doi : 10.1006/dbio.1999.9427 . PMID 10525332.
- ^ Afzelius, BA (1961). "La estructura fina de los cilios de las placas natatorias de los ctenóforos". Revista de citología biofísica y bioquímica . 9 (2): 383–94. doi :10.1083/jcb.9.2.383. PMC 2224992 . PMID 13681575.
- ^ Kreps, TA; Purcell, JE y Heidelberg, KB (junio de 1997). "Escape del ctenóforo Mnemiopsis leidyi del depredador scyphomedusa Chrysaora quinquecirrha ". Biología Marina . 128 (3): 441–446. Código Bib : 1997MarBi.128..441K. doi :10.1007/s002270050110. S2CID 32975367.
- ^ Mills, Claudia E. (febrero de 1984). "La densidad se altera en hidromedusas y ctenóforos en respuesta a cambios en la salinidad". The Biological Bulletin . 166 (1): 206–215. doi :10.2307/1541442. JSTOR 1541442.
- ^ Las medusas peine, que parecen extraterrestres, tienen un sistema nervioso como nunca antes se había visto.
- ^ La medusa con sistema nervioso que está causando escalofríos en la comunidad científica
- ^ ab Burkhardt, Pawel; Colgren, Jeffrey; Medhus, Astrid; Digel, Leónidas; Naumann, Benjamín; Soto-Ángel, Joan; et al. (20 de abril de 2023). "La red nerviosa sincitial en un ctenóforo aporta información sobre la evolución de los sistemas nerviosos". Ciencia . 380 (6642): 293–297. Código Bib : 2023 Ciencia... 380.. 293B. doi : 10.1126/science.ade5645. PMID 37079688. S2CID 258239574 . Consultado el 24 de abril de 2023 .
- ^ Parry, Luke A.; Lerosey-Aubril, Rudy; Weaver, James C.; Ortega-Hernández, Javier (2021). "Las medusas peine del Cámbrico de Utah iluminan la evolución temprana de los sistemas nervioso y sensorial en los ctenóforos". iScience . 24 (9): 102943. Bibcode :2021iSci...24j2943P. doi : 10.1016/j.isci.2021.102943 . PMC 8426560 . PMID 34522849.
- ^ Pennisi, Elizabeth (10 de enero de 2019). "Los tentáculos pegajosos de las medusas peine pueden haber revelado cuándo evolucionaron por primera vez las células nerviosas". Science . doi :10.1126/science.aaw6288. S2CID 92852830.
- ^¿ El sistema nervioso ctenóforo evolucionó de forma independiente?
- ^ Marlow, Heather; Tosches, Maria Antonietta; Tomer, Raju; Steinmetz, Patrick R.; Lauri, Antonella; Larsson, Tomas; Arendt, Detlev (29 de enero de 2014). "El patrón del cuerpo larvario y los órganos apicales se conservan en la evolución animal". BMC Biology . 12 (1): 7. doi : 10.1186/1741-7007-12-7 . PMC 3939940 . PMID 24476105.
- ^ Nielsen, Claus (15 de febrero de 2015). "Sistemas nerviosos larvarios: larvas verdaderas y adultos precoces". Journal of Experimental Biology . 218 (4): 629–636. doi : 10.1242/jeb.109603 . PMID 25696826. S2CID 3151957.
- ^ Nielsen, Claus (julio de 2019). «Evolución animal temprana: la visión de un morfólogo». Royal Society Open Science . 6 (7): 190638. Bibcode :2019RSOS....690638N. doi :10.1098/rsos.190638. PMC 6689584 . PMID 31417759.
- ^ Fox, Douglas (1 de agosto de 2017). «Alienígenas entre nosotros». Aeon . Consultado el 1 de agosto de 2017 .
- ^ Hewitt, Olivia H.; Degnan, Sandie M. (13 de febrero de 2023). "Las enzimas antioxidantes que atacan al peróxido de hidrógeno se conservan en todo el reino animal, desde las esponjas hasta los mamíferos". Scientific Reports . 13 (1): 2510. Bibcode :2023NatSR..13.2510H. doi :10.1038/s41598-023-29304-6. PMC 9925728 . PMID 36781921. S2CID 256811787.
- ^ Norekian, Tigran P.; Moroz, Leonid L. (15 de agosto de 2019). "Sistema neural y diversidad de receptores en el ctenóforo Beroe abyssicola". Journal of Comparative Neurology . 527 (12): 1986–2008. doi : 10.1002/cne.24633 . PMID 30632608.
- ^ Simmons, David K.; Martindale, Mark Q. (2015). "Ctenophora". Estructura y evolución de los sistemas nerviosos de los invertebrados . Oxford University Press. págs. 48-55. doi :10.1093/acprof:oso/9780199682201.003.0006. ISBN . 9780199682201.
- ^ Jákely, Gáspár; Paps, Jordi; Nielsen, Claus (2015). "La posición filogenética de los ctenóforos y el origen de los sistemas nerviosos". EvoDevo . 6 (1): 1. doi : 10.1186/2041-9139-6-1 . PMC 4406211 . PMID 25905000.
- ^ Cinco tipos de coloblastos en un ctenóforo cídipido, Minictena luteola Carré y Carré: un estudio ultraestructural e interpretación citológica
- ^ Caracterización de la biodiversidad funcional en el filo Ctenophora mediante mediciones fisiológicas
- ^ Mills, CE; Miller, RL (febrero de 1984). "Ingestión de una medusa ( Aegina citrea ) por el ctenóforo que contiene nematocistos ( Haeckelia rubra , anteriormente Euchlora rubra ): implicaciones filogenéticas". Biología marina . 78 (2): 215–221. Bibcode :1984MarBi..78..215M. doi :10.1007/BF00394704. S2CID 17714037.
- ^ Mackie, VAMOS; Molinos, CE; Singla, CL (marzo de 1988). "Estructura y función de la tentilla prensil de Euplokamis (Ctenophora, Cydippida)". Zoomorfología . 107 (6): 319–337. doi :10.1007/BF00312216. S2CID 317017.
- ^ ab Craig, CL; Okubo, A. (abril de 1990). "Restricciones físicas en la evolución del tamaño y la forma de los ctenóforos". Ecología evolutiva . 4 (2): 115–129. Bibcode :1990EvEco...4..115C. doi :10.1007/BF02270909. S2CID 24584197.
- ^ ab Tamm, Sidney L. (1973). "Mecanismos de coordinación ciliar en ctenóforos". Journal of Experimental Biology . 59 : 231–245. doi :10.1242/jeb.59.1.231.
- ^ abcdef Haddock, SHD (diciembre de 2007). "Comportamiento alimentario comparativo de los ctenóforos planctónicos". Biología comparativa e integradora . 47 (6): 847–853. doi :10.1093/icb/icm088. PMID 21669763.
- ^ Tamm, SL; Tamm, S. (1985). "Visualización de los cambios en la configuración de la punta ciliar causados por el desplazamiento deslizante de los microtúbulos en los macrocilios del ctenóforo Beroe ". Journal of Cell Science . 79 : 161–179. doi :10.1242/jcs.79.1.161. PMID 3914479.
- ^ Tamm, Sidney L.; Tamm, Signhild (1991). "La adhesión epitelial reversible cierra la boca de Beroe, una medusa marina carnívora". Boletín biológico . 181 (3): 463–473. doi :10.2307/1542367. JSTOR 1542367. PMID 29304670.
- ^ Gibbons, Mark J.; Richardson, Anthony J.; V. Angel, Martin; Buecher, Emmanuelle; Esnal, Graciela; Fernandez Alamo, Maria A.; et al. (junio de 2005). "Qué determina la probabilidad de descubrimiento de especies en el holozooplancton marino: ¿es importante el tamaño, la distribución o la profundidad?". Oikos . 109 (3): 567–576. Bibcode :2005Oikos.109..567G. doi :10.1111/j.0030-1299.2005.13754.x.
- ^ Wrobel, David; Mills, Claudia (2003) [1998]. Invertebrados pelágicos de la costa del Pacífico: una guía de los animales gelatinosos más comunes. Sea Challengers y Monterey Bay Aquarium. pp. 108. ISBN 978-0-930118-23-5.
- ^ Martindale, MQ (diciembre de 1986). "La ontogenia y el mantenimiento de las propiedades de simetría adulta en el ctenóforo, Mnemiopsis mccradyi ". Biología del desarrollo . 118 (2): 556–576. doi :10.1016/0012-1606(86)90026-6. PMID 2878844.
- ^ Integración fisiológica rápida de ctenóforos fusionados
- ^ Sasson, Daniel A.; Ryan, Joseph F. (diciembre de 2017). "Una reconstrucción de los modos sexuales a lo largo de la evolución animal". BMC Evolutionary Biology . 17 (1): 242. Bibcode :2017BMCEE..17..242S. doi : 10.1186/s12862-017-1071-3 . PMC 5717846 . PMID 29207942.
- ^ Edgar, Allison; Ponciano, José Miguel; Martindale, Mark Q. (2022). "Los ctenóforos son desarrolladores directos que se reproducen continuamente comenzando muy temprano después de la eclosión". Actas de la Academia Nacional de Ciencias de los EE. UU . . 119 (18): e2122052119. Bibcode :2022PNAS..11922052E. doi : 10.1073/pnas.2122052119 . PMC 9170174 . PMID 35476523.
- ^ Harbison, GR; Miller, RL (febrero de 1986). "No todos los ctenóforos son hermafroditas. Estudios sobre la sistemática, distribución, sexualidad y desarrollo de dos especies de Ocyropsis". Biología marina . 90 (3): 413–424. Bibcode :1986MarBi..90..413H. doi :10.1007/bf00428565. S2CID 83954780.
- ^ Reitzel, AM; Pang, K.; Martindale, MQ (2016). "Expresión del desarrollo de genes relacionados con la 'línea germinal' y la 'determinación sexual' en el ctenóforo Mnemiopsis leidyi". EvoDevo . 7 : 17. doi : 10.1186/s13227-016-0051-9 . PMC 4971632 . PMID 27489613.
- ^ Jaspers, C.; Haraldsson, M.; Bolte, S.; Reusch, TB; Thygesen, UH; Kiørboe, T. (2012). "La población de ctenóforos se recluta completamente a través de la reproducción larvaria en el mar Báltico central". Biology Letters . 8 (5): 809–12. doi :10.1098/rsbl.2012.0163. PMC 3440961 . PMID 22535640.
- ^ Moroz, Leonid; Mukherjee, Krishanu; Romanova, Daria (2023). "Señalización de óxido nítrico en ctenóforos". Portada. Neurosci . 17 : 1125433. doi : 10.3389/fnins.2023.1125433 . PMC 10073611 . PMID 37034176.
- ^ Esta medusa parecida a Benjamin Button puede envejecer en reversa, de adulta a juvenil
- ^ abcd Mills, CE (mayo de 2007). «Phylum Ctenophora: lista de todos los nombres científicos válidos» . Consultado el 10 de febrero de 2009 .
- ^ Welch, Victoria; Vigneron, Jean Pol; Lousse, Virginie; Parker, Andrew (14 de abril de 2006). "Propiedades ópticas del órgano iridiscente de la medusa peine Beroë cucumis (Ctenophora)". Physical Review E . 73 (4): 041916. Bibcode :2006PhRvE..73d1916W. doi :10.1103/PhysRevE.73.041916. PMID 16711845.
- ^ Haddock, SHD; Case, JF (diciembre de 1995). "No todos los ctenóforos son bioluminiscentes: Pleurobrachia". Boletín biológico . 189 (3): 356–362. doi :10.2307/1542153. JSTOR 1542153. PMID 29244577.
- ^ Abadejo, SHD; Caso, JF (8 de abril de 1999). "Espectros de bioluminiscencia del zooplancton gelatinoso de aguas poco profundas y profundas: ctenóforos, medusas y sifonóforos". Biología Marina . 133 (3): 571–582. Código Bib : 1999MarBi.133..571H. doi :10.1007/s002270050497. S2CID 14523078.
- ^ Schnitzler, Christine E.; Pang, Kevin; Powers, Meghan L.; Reitzel, Adam M.; Ryan, Joseph F.; Simmons, David; et al. (2012). "Organización genómica, evolución y expresión de genes de fotoproteína y opsina en Mnemiopsis leidyi: una nueva visión de los fotócitos ctenóforos". BMC Biology . 10 (1): 107. doi : 10.1186/1741-7007-10-107 . PMC 3570280 . PMID 23259493.
- ^ Winnikoff, Jacob R.; Haddock, Steven HD; Budin, Itay (1 de noviembre de 2021). "Adaptaciones de ácidos grasos específicas de la profundidad y la temperatura en ctenóforos de hábitats extremos". Journal of Experimental Biology . 224 (21): jeb242800. doi :10.1242/jeb.242800. PMC 8627573 . PMID 34676421.
- ^ el-Shabrawy, Gamal; Dumont, Henri (2016). "Primer registro de un ctenóforo en lagos: la medusa peine Mnemiopsis leidyi (A. Agassiz, 1865) invade el Fayum, Egipto". BioInvasions Records . 5 (1): 21–24. doi : 10.3391/bir.2016.5.1.04 . S2CID 90377075.
- ^ Mohammed-Geba, Khaled; Sheir, Sherin; Aguilar, Roberto; Ogburn, Mateo; Hines, Anson; Khalafallah, Hussain; et al. (2019). "Confirmación molecular y morfológica de un isópodo americano invasor; Livoneca redmanii (Leach, 1818), desde la región mediterránea hasta el lago Qaroun, Egipto" (PDF) . Revista egipcia de biología y pesca acuáticas . 23 (4): 251–273. doi :10.21608/ejabf.2019.54062.
- ^ Reeve, MR; Walter, MA; Ikeda, T. (julio de 1978). "Estudios de laboratorio sobre la ingestión y utilización de alimentos en ctenóforos lobulados y tentaculados 1: Utilización de alimentos por ctenóforos". Limnología y Oceanografía . 23 (4): 740–751. doi : 10.4319/lo.1978.23.4.0740 .
- ^ Costello, JH; Coverdale, R. (octubre de 1998). "Alimentación planctónica y significado evolutivo del plan corporal lobulado dentro de los Ctenophora". The Biological Bulletin . 195 (2): 247–248. doi :10.2307/1542863. JSTOR 1542863. PMID 28570175.
- ^ Bos, AR; Cruz-Rivera, E.; Sanad, AM (2016). "Los peces herbívoros Siganus rivulatus (Siganidae) y Zebrasoma desjardinii (Acanthuridae) se alimentan de Ctenophora y Scyphozoa en el Mar Rojo". Biodiversidad Marina . 47 : 243–246. doi :10.1007/s12526-016-0454-9. S2CID 24694789.
- ^ Arai, Mary Needler (junio de 2005). "Depredación de celentéreos pelágicos: una revisión". Revista de la Asociación de Biología Marina del Reino Unido . 85 (3): 523–536. Bibcode :2005JMBUK..85..523A. doi :10.1017/S0025315405011458. S2CID 86663092.
- ^ ab Chandy, Shonali T.; Greene, Charles H. (julio de 1995). "Estimación del impacto depredador del zooplancton gelatinoso". Limnología y Oceanografía . 40 (5): 947–955. Bibcode :1995LimOc..40..947C. doi : 10.4319/lo.1995.40.5.0947 .
- ^ Field, Christopher B.; Behrenfeld, Michael J.; Randerson, James T.; Falkowski, Paul (10 de julio de 1998). "Producción primaria de la biosfera: integración de componentes terrestres y oceánicos". Science . 281 (5374): 237–240. Bibcode :1998Sci...281..237F. doi :10.1126/science.281.5374.237. PMID 9657713.
- ^ abc Purcell, Jennifer E.; Shiganova, Tamara A.; Decker, Mary Beth; Houde, Edward D. (2001). "El ctenóforo Mnemiopsis en hábitats nativos y exóticos: estuarios de Estados Unidos versus la cuenca del Mar Negro". Hydrobiologia . 451 (1–3): 145–176. doi :10.1023/A:1011826618539. S2CID 23336715.
- ^ ab Oguz, T.; Fach, B. y Salihoglu, B. (diciembre de 2008). "Dinámica de invasión del ctenóforo exótico Mnemiopsis leidyi y su impacto en el colapso de la anchoa en el Mar Negro". Journal of Plankton Research . 30 (12): 1385–1397. doi : 10.1093/plankt/fbn094 .
- ^ Bayha, KM; Harbison, R.; McDonald, JH; Gaffney, PM (2004). "Investigación preliminar sobre la sistemática molecular del ctenóforo invasivo Beroe ovata ". En Dumont, H.; Shiganova, TA; Niermann, U. (eds.). Invasiones acuáticas en los mares Negro, Caspio y Mediterráneo . Serie de Ciencias IV de la OTAN: Ciencias de la Tierra y el Medio Ambiente. vol. 35. Springer Países Bajos. págs. 167-175. doi :10.1007/1-4020-2152-6_7. ISBN 978-1-4020-1866-4.
- ^ Kamburska, L. (2006). "Efectos de Beroe cf ovata en el zooplancton gelatinoso y de otro tipo a lo largo de la costa búlgara del Mar Negro". En Dumont, H.; Shiganova, TA; Niermann, U. (eds.). Invasiones acuáticas en los mares Negro, Caspio y Mediterráneo . Serie de Ciencias de la OTAN IV: Ciencias de la Tierra y del Medio Ambiente. Vol. 35. Springer Netherlands. págs. 137–154. doi :10.1007/1-4020-2152-6_5. ISBN 978-1-4020-1866-4.
- ^ "Las neuronas de la medusa peine provocan un debate sobre la evolución". Quanta Magazine . 25 de marzo de 2015 . Consultado el 12 de junio de 2015 .
- ^ Whelan, NV; Kocot, KM; Moroz, TP; Mukherjee, K.; Williams, P.; Paulay, G.; et al. (2017). "Relaciones de los ctenóforos y su ubicación como grupo hermano de todos los demás animales". Nature Ecology and Evolution . 1 (11): 1737–1746. Bibcode :2017NatEE...1.1737W. doi :10.1038/s41559-017-0331-3. PMC 56641 . PMID 28993654.
- ^ Conway Morris, S. (2003). "La "explosión" cámbrica de metazoos y biología molecular: ¿Estaría Darwin satisfecho?" (PDF) . Revista internacional de biología del desarrollo . 47 (7–8): 505–515. PMID 14756326. Archivado desde el original (PDF) el 24 de diciembre de 2009. Consultado el 14 de febrero de 2009 .
- ^ Klug, Christian; Kerr, Johanne; Lee, Michael SY; Cloutier, Richard (2021). "Un tallo ctenóforo superviviente tardío del Devónico tardío de Miguasha (Canadá)". Scientific Reports . 11 (1): 19039. Bibcode :2021NatSR..1119039K. doi :10.1038/s41598-021-98362-5. PMC 8463547 . PMID 34561497.
- ^ Maxmen, Amy (7 de septiembre de 2011). «Una antigua medusa marina sacude el árbol evolutivo de los animales». Scientific American . Consultado el 21 de junio de 2018 .
- ^ Shu, D.-G.; Conway Morris, S. ; Han, J.; Li, Y.; Zhang, XL; Hua, H.; et al. (5 de mayo de 2006). "Vendobiontes del Cámbrico Inferior de China y evolución temprana de diploblastos". Science . 312 (5774): 731–734. Bibcode :2006Sci...312..731S. doi :10.1126/science.1124565. PMID 16675697. S2CID 1235914.
- ^ Zhao, Yang; Vinther, Jakob; Parry, Luke A.; Wei, Fan; Green, Emily; Pisani, Davide; et al. (abril de 2019). "Ctenóforos sésiles del grupo troncal cámbrico que se alimentan por suspensión y evolución del plan corporal de las medusas peine". Current Biology . 29 (7): 1112–1125.e2. doi : 10.1016/j.cub.2019.02.036 . PMID 30905603.
- ^ Zhao, Yang; Hou, Xian-guang; Cong, Pei-yun (enero de 2023). "Naturaleza tentacular de la 'columna' de la diploblástica Xianguangia sinica del Cámbrico". Revista de Paleontología Sistemática . 21 (1). Código Bibliográfico :2023JSPal..2115787Z. doi : 10.1080/14772019.2023.2215787 . ISSN 1477-2019.
- ^ Mindy, Weisberger (10 de julio de 2015). "Las medusas antiguas tenían esqueletos espinosos, no tentáculos". livescience.com .
- ^ Conway Morris, Simon ; Simonetta, Alberto M., eds. (1991). La evolución temprana de los metazoos y la importancia de los taxones problemáticos . Cambridge University Press. pág. 308. ISBN 978-0-521-11158-4.
- ^ Nielsen, Claus; Scharff, Nikolaj; Eibye-Jacobsen, Danny (abril de 1996). "Análisis cladísticos del reino animal". Revista biológica de la Sociedad Linneana . 57 (4): 385–410. Bibcode :1996BJLS...57..385N. doi : 10.1006/bijl.1996.0023 .
- ^ Leuckart, Rudolf (1923). Ueber die Morphologie und die Verwandtschaftsverhältnisse der wirbellosen thiere. Ein Beitrag zur Charakteristik und Classification der thierischen Formen (en alemán). ISBN 978-1-245-56027-6.
- ^ Haeckel, Ernst (1896). Systematische Phylogenie der Wirbellosen Thière . Invertebrata (en alemán). vol. Parte 2: Des Entwurfs Einer Systematischen Stammesgeschichte. ISBN 978-1-120-86850-3.
- ^ Hyman, Libbie Henrietta (1940). Los invertebrados . Vol. I Protozoos a Ctenophora. McGraw Hill. ISBN 978-0-07-031660-7.
- ^ Philippe, H.; Derelle, R.; Lopez, P.; Pick, K.; Borchiellini, C.; Boury-Esnault, N.; et al. (28 de abril de 2009). "La filogenómica revive las visiones tradicionales sobre las relaciones profundas entre animales". Current Biology . 19 (8): 706–712. Bibcode :2009CBio...19..706P. doi : 10.1016/j.cub.2009.02.052 . PMID 19345102. S2CID 15282843.
- ^ Wallberg, A.; Thollesson, M.; Farris, JS; Jondelius, U. (diciembre de 2004). "La posición filogenética de las medusas peine (Ctenophora) y la importancia del muestreo taxonómico". Cladistics . 20 (6): 558–578. doi : 10.1111/j.1096-0031.2004.00041.x . PMID 34892961. S2CID 86185156.
- ^ Collins, AG (2002). "Filogenia de Medusozoa y la evolución de los ciclos de vida de los cnidarios". Journal of Evolutionary Biology . 15 (3): 418–432. doi : 10.1046/j.1420-9101.2002.00403.x . S2CID 11108911.
- ^ ab Pick, KS; Philippe, H.; Schreiber, F.; Erpenbeck, D.; Jackson, DJ; Wrede, P.; et al. (septiembre de 2010). "El muestreo de taxones filogenómicos mejorado afecta notablemente las relaciones no bilaterales". Biología molecular y evolución . 27 (9): 1983–1987. doi :10.1093/molbev/msq089. PMC 2922619 . PMID 20378579.
- ^ Hejnol, A.; Obst, M.; Stamatakis, A.; Ott, M.; Rouse, GW; Edgecombe, GD; et al. (22 de diciembre de 2009). "Evaluación de la raíz de animales bilaterales con métodos filogenómicos escalables". Actas de la Royal Society B: Biological Sciences . 276 (1677): 4261–4270. doi :10.1098/rspb.2009.0896. PMC 2817096 . PMID 19759036.
- ^ Ryan, JF; Pang, K.; Programa de secuenciación comparativa; Mullikin, JC; Martindale, MQ; Baxevanis, AD; Programa de secuenciación comparativa del NISC (2010). "El complemento de homeodominio del ctenóforo Mnemiopsis leidyi sugiere que Ctenophora y Porifera divergieron antes que ParaHoxozoa". EvoDevo . 1 (1): 9. doi : 10.1186/2041-9139-1-9 . PMC 2959044 . PMID 20920347.
- ^ Reitzel, AM; Pang, K.; Ryan, JF; Mullikin, JC; Martindale, MQ; Baxevanis, AD; Tarrant, AM (2011). "Los receptores nucleares del ctenóforo Mnemiopsis leidyi carecen de un dominio de unión al ADN de dedo de zinc: ¿pérdida específica de linaje o condición ancestral en el surgimiento de la superfamilia de receptores nucleares?". EvoDevo . 2 (1): 3. doi : 10.1186/2041-9139-2-3 . PMC 3038971 . PMID 21291545.
- ^ Pang, K.; Ryan, JF; NISC Comparative Sequencing Program; Mullikin, JC; Baxevanis, AD; Martindale, MQ (2010). "Información genómica sobre la señalización de Wnt en un metazoo divergente temprano, el ctenóforo Mnemiopsis leidyi". EvoDevo . 1 (1): 10. doi : 10.1186/2041-9139-1-10 . PMC 2959043 . PMID 20920349.
- ^ Liebeskind, BJ; Hillis, DM; Zakon, HH (2011). "La evolución de los canales de sodio es anterior al origen de los sistemas nerviosos en los animales". Actas de la Academia Nacional de Ciencias de los Estados Unidos . 108 (22): 9154–9159. Bibcode :2011PNAS..108.9154L. doi : 10.1073/pnas.1106363108 . PMC 3107268 . PMID 21576472.
- ^ ab Ryan, JF; Pang, K.; Schnitzler, CE; Nguyen, A.-D.; Moreland, RT; Simmons, DK; et al. (13 de diciembre de 2013). "El genoma del ctenóforo Mnemiopsis leidyi y sus implicaciones para la evolución del tipo celular". Science . 342 (6164): 1242592. doi :10.1126/science.1242592. PMC 3920664 . PMID 24337300.
- ^ Whelan, Nathan V.; Kocot, Kevin M.; Moroz, Leonid L.; Halanych, Kenneth M. (5 de mayo de 2015). "Error, señal y la ubicación de la hermana de Ctenophora con respecto a todos los demás animales". Actas de la Academia Nacional de Ciencias de los Estados Unidos . 112 (18): 5773–5778. Bibcode :2015PNAS..112.5773W. doi : 10.1073/pnas.1503453112 . PMC 4426464 . PMID 25902535.
- ^ Borowiec, Marek L.; Lee, Ernest K.; Chiu, Joanna C.; Plachetzki, David C. (diciembre de 2015). "La extracción de señales filogenéticas y la consideración del sesgo en los conjuntos de datos del genoma completo respaldan a Ctenophora como hermana del resto de los metazoos". BMC Genomics . 16 (1): 987. doi : 10.1186/s12864-015-2146-4 . PMC 4657218 . PMID 26596625.
- ^ Philippe, Hervé; Derelle, Romain; Lopez, Philippe; Pick, Kerstin; Borchiellini, Carole; Boury-Esnault, Nicole; et al. (abril de 2009). "La filogenómica revive las opiniones tradicionales sobre las relaciones profundas entre animales". Current Biology . 19 (8): 706–712. Bibcode :2009CBio...19..706P. doi : 10.1016/j.cub.2009.02.052 . PMID 19345102. S2CID 15282843.
- ^ Nosenko, Tetyana; Schreiber, Fabián; Adamska, Maja; Adamski, Marcin; Eitel, Michael; Hammel, Jörg; et al. (Abril de 2013). "Filogenia profunda de los metazoos: cuando diferentes genes cuentan historias diferentes". Filogenética molecular y evolución . 67 (1): 223–233. Código Bib : 2013 MolPE..67..223N. doi :10.1016/j.ympev.2013.01.010. PMID 23353073.
- ^ Pisani, Davide; Pett, Walker; Dohrmann, Martin; Feuda, Roberto; Rota-Stabelli, Omar; Philippe, Hervé; et al. (15 de diciembre de 2015). "Los datos genómicos no respaldan que las medusas peine sean el grupo hermano de todos los demás animales". Actas de la Academia Nacional de Ciencias de los Estados Unidos . 112 (50): 15402–15407. Bibcode :2015PNAS..11215402P. doi : 10.1073/pnas.1518127112 . PMC 4687580 . PMID 26621703.
- ^ Kapli, Paschalia; Telford, Maximilian J. (11 de diciembre de 2020). "La asimetría dependiente de la topología en los errores sistemáticos afecta la ubicación filogenética de Ctenophora y Xenacoelomorpha". Science Advances . 6 (10): eabc5162. Bibcode :2020SciA....6.5162K. doi :10.1126/sciadv.abc5162. PMC 7732190 . PMID 33310849.
- ^ Christianson, LM; Johnson, SB; Schultz, DT; Haddock, SH (2021). "Diversidad oculta de Ctenophora revelada por nuevos cebadores y secuencias COI mitocondriales". Recursos de ecología molecular . 22 (1): 283–294. doi :10.1111/1755-0998.13459. PMC 9290464 . PMID 34224654.
- ^ Kohn, AB; Citarella, MR; Kocot, KM; Bobkova, YV; Halanych, KM; Moroz, LL (2011). "Evolución rápida del genoma mitocondrial compacto e inusual en el ctenóforo, Pleurobrachia bachei". Filogenética molecular y evolución . 63 (1): 203–207. doi :10.1016/j.ympev.2011.12.009. PMC 4024468 . PMID 22201557.
- ^ Bakshani, Cassie R.; Morales-García, Ana L.; Althaus, Mike; Wilcox, Matthew D.; Pearson, Jeffrey P.; Bythell, John C.; Burgess, J. Grant (4 de julio de 2018). "Conservación evolutiva de la función antimicrobiana del moco: una primera defensa contra la infección". NPJ Biofilms and Microbiomes . 4 (1): 14. doi :10.1038/s41522-018-0057-2. PMC 6031612 . PMID 30002868.
- ^ Okubo, Tomomi (8 de agosto de 2022). "En el cerebro de las medusas peine: los científicos exploran la evolución de las neuronas". neurosciencenews.com (Comunicado de prensa). Okinawa, JA: Instituto de Ciencia y Tecnología de Okinawa .
Un estudio de las medusas peine revela el tipo de mensajero que probablemente funcionó en el sistema nervioso ancestral.
— cita artículo de revista: - ^ Mills, DB; Francis, WR; Vargas, S; Larsen, M; Elemans, CP; Canfield, DE; Wörheide, G (2018). "El último ancestro común de los animales carecía de la vía HIF y respiraba en ambientes con poco oxígeno". eLife . 7 . doi : 10.7554/eLife.31176 . PMC 5800844 . PMID 29402379.
- ^ Gauberg, Julia; Abdallah, Salsabil; Elkhatib, Wassim; Harracksingh, Alicia N.; Piekut, Thomas; Stanley, Elise F.; Senatore, Adriano (25 de diciembre de 2020). "Características biofísicas conservadas del homólogo del canal Ca2+ presináptico CaV2 del animal de divergencia temprana Trichoplax adhaerens". The Journal of Biological Chemistry . 295 (52): 18553–18578. doi : 10.1074/jbc.RA120.015725 . PMC 7939481 . PMID 33097592.
- ^ Spradling, Allan (14 de mayo de 2012). «Relaciones de los primeros metazoos». Evolución y desarrollo (PDF) (diapositivas de la presentación). Programa de investigación Evo-Devo. Washington, DC: Carnagie Science . p. 38. Archivado desde el original (PDF) el 2 de marzo de 2014. Consultado el 8 de octubre de 2024 en carnegiescience.edu.— Introducción técnica y revisión de la investigación genómica, la evolución y la taxonomía.
- ^ Hiebert, Laurel S.; Maslakova, Svetlana A. (2015). "Los genes Hox modelan el eje anteroposterior del juvenil pero no de la larva en un invertebrado de desarrollo indirecto máximo, Micrura alaskensis (Nemertea)". BMC Biology . 13 : 23. doi : 10.1186/s12915-015-0133-5 . PMC 4426647 . PMID 25888821.
- ^ Gąsiorowski, Ludwik; Hejnol, Andreas (2019). "Expresión del gen Hox durante el desarrollo del forónido Phoronopsis harmeri". EvoDevo . 11 : 2. bioRxiv 10.1101/799056 . doi : 10.1186/s13227-020-0148-z . PMC 7011278 . PMID 32064072. S2CID 208578827.
- ^ Señalización independiente en forma de radiación de innexina en ctenóforos
- ^ ab Podar, Mircea; Abadejo, Steven HD; Sogin, Mitchell L.; Harbison, G. Richard (noviembre de 2001). "Un marco filogenético molecular para el filo Ctenophora utilizando genes de ARNr 18S". Filogenética molecular y evolución . 21 (2): 218–230. Código Bib : 2001MolPE..21..218P. CiteSeerX 10.1.1.384.6705 . doi :10.1006/mpev.2001.1036. PMID 11697917.
- ^ Harbison, GR (1985). "Sobre la clasificación y evolución de los ctenóforos". En Conway Morris, S .; George, JD; Gibson, R.; Platt, HM (eds.). Los orígenes y relaciones de los invertebrados inferiores . Clarendon Press. págs. 78-100. ISBN 978-0-19-857181-0.
- ^ Simion, Paul; Bekkouche, Nicolas; Jager, Muriel; Quéinnec, Eric; Manuel, Michaël (2015). "Explorando el potencial de las subunidades pequeñas de ARN y las secuencias ITS para resolver las relaciones filogenéticas dentro del filo Ctenophora". Zoología . 118 (2): 102–114. Bibcode :2015Zool..118..102S. doi :10.1016/j.zool.2014.06.004. PMID 25440713.
Lectura adicional
- Barnes, RSK; Calow, P.; Olive, PJW; Golding, DW; Spicer, JI (2001). Los invertebrados: una síntesis (3.ª ed.). Blackwell. § 3.4.3, pág. 63. ISBN 0-632-04761-5.
- Brusca, RC; Brusca, GJ (2003). Invertebrados (2ª ed.). Asociados Sinauer. cap. 9, pág. 269.ISBN 0-87893-097-3.
- Moore, J. (2001). Introducción a los invertebrados . Cambridge University Press. § 5.4, pág. 65. ISBN 0-521-77914-6.
- Schäfer, W. (1996). "Ctenófora, Rippenquallen". En Westheide, W.; Rieger, R. (eds.). Spezielle Zoología . vol. 1. Stuttgart, Alemania: Gustav Fischer Verlag.
- Wenzel, Bruno (1958). Glastiere des Meeres. Rippenquallen (Acnidaria) . ISBN 3-7403-0189-9.
- Shasha, Mark (1992). La noche de las medusas lunares . Simon & Schuster. ISBN 0-671-77565-0.
- Fox, Douglas (agosto de 2017). “Alienígenas entre nosotros: lo que el ctenóforo dice sobre la evolución de la inteligencia”. Aeon . Londres, Reino Unido / Nueva York, NY / Melbourne, AU: Aeon Media Group – vía Aeon.co.
El cerebro del ctenóforo sugiere que, si la evolución comenzara de nuevo, la inteligencia resurgiría porque la naturaleza se repite.
— en parte una revisión de la investigación hasta Moroz et al. (2014)
Enlaces externos
 Definiciones de Wikcionario
Definiciones de Wikcionario- Medios de comunicación de Commons
- Taxones de Wikispecies
- Datos de Wikidata
- Crónicas del plancton (vídeo). Archivado desde el original el 12 de enero de 2012.— Cortometrajes documentales y fotografías
- "Medusas y medusas peine". Instituto Smithsoniano . 30 de abril de 2018 – vía ocean.si.edu.— descripción general en el portal Smithsonian Ocean
- "Ctenóforos del Canal São Sebastião, Brasil". Archivado desde el original el 4 de febrero de 2012 - vía usp.br.
- Vídeo de ctenóforos en el Zoológico Nacional de Washington DC (video) – vía YouTube .
- "Los biólogos evolucionistas han reorganizado las ramas del árbol de la vida animal". sciencedaily.com (nota de prensa). Marzo de 2008.
- Ctenophora. Guía del zooplancton marino del sudeste de Australia (ficha técnica). Instituto de Acuicultura y Pesca de Tasmania / División Antártica Australiana. Departamento de Medio Ambiente, Agua, Patrimonio y Artes del Gobierno de Australia. Junio de 2008. Archivado desde el original el 20 de julio de 2008. Consultado el 8 de octubre de 2024 en tafi.org.au.
{{cite report}}: CS1 maint: URL no apta ( enlace )— incluye imágenes - La conexión de gelatina. imagequest3d.com (video mixto e imágenes fijas).— imágenes impactantes, incluido un espécimen de Beroe atacando a otro ctenóforo
- «En busca de los primeros animales». National Geographic . 12 de diciembre de 2013. Archivado desde el original el 12 de diciembre de 2013.