Flujo de energía (ecología)
{kind=link}

{kind=link}
El flujo de energía es el flujo de energía a través de los seres vivos dentro de un ecosistema . [1] Todos los organismos vivos pueden organizarse en productores y consumidores , y esos productores y consumidores pueden organizarse a su vez en una cadena alimentaria . [2] [3] Cada uno de los niveles dentro de la cadena alimentaria es un nivel trófico . [1] Para mostrar de manera más eficiente la cantidad de organismos en cada nivel trófico, estas cadenas alimentarias se organizan en pirámides tróficas . [1] Las flechas en la cadena alimentaria muestran que el flujo de energía es unidireccional, con la punta de una flecha indicando la dirección del flujo de energía; la energía se pierde como calor en cada paso del camino. [2] [3]
El flujo unidireccional de energía y la pérdida sucesiva de energía a medida que viaja a lo largo de la cadena alimentaria son patrones en el flujo de energía que están regidos por la termodinámica , que es la teoría del intercambio de energía entre sistemas. [4] [5] La dinámica trófica se relaciona con la termodinámica porque se ocupa de la transferencia y transformación de energía (que se origina externamente desde el sol a través de la radiación solar) hacia y entre organismos. [1]
La energía y el ciclo del carbono
{kind=link}

100×10 15 gramos de carbono/año fijados por los organismos fotosintéticos, lo que equivale a 4×10 18 kJ/año = 4×10 21 J/año de energía libre.La respiración celular es la reacción inversa, en la que se absorbe energía de las plantas y se desprende dióxido de carbono y agua. El dióxido de carbono y el agua producidos se pueden reciclar nuevamente en las plantas.
El primer paso en la energía es la fotosíntesis , en la que el agua y el dióxido de carbono del aire se absorben con la energía del sol y se convierten en oxígeno y glucosa. [7] La respiración celular es la reacción inversa, en la que se absorben oxígeno y azúcar y se libera energía a medida que se convierten nuevamente en dióxido de carbono y agua. El dióxido de carbono y el agua producidos por la respiración se pueden reciclar nuevamente en las plantas.
La pérdida de energía se puede medir por la eficiencia (cuánta energía llega al siguiente nivel) o por la biomasa (cuánto material vivo existe en esos niveles en un momento dado, medido por la cantidad de cultivos en pie). [1] De toda la productividad primaria neta en el nivel trófico de productores, en general solo el 10% va al siguiente nivel, los consumidores primarios, luego solo el 10% de ese 10% pasa al siguiente nivel trófico, y así sucesivamente hasta la pirámide alimentaria. [1] La eficiencia ecológica puede estar entre el 5% y el 20% dependiendo de qué tan eficiente o ineficiente sea ese ecosistema. [8] [1] Esta disminución en la eficiencia ocurre porque los organismos necesitan realizar respiración celular para sobrevivir, y la energía se pierde en forma de calor cuando se realiza la respiración celular. [1] Esa es también la razón por la que hay menos consumidores terciarios que productores. [1]
Producción primaria
{kind=link}

Un productor es cualquier organismo que realiza la fotosíntesis . [9] Los productores son importantes porque convierten la energía del sol en una forma química de energía almacenable y utilizable, la glucosa, [1] así como el oxígeno. Los propios productores pueden utilizar la energía almacenada en la glucosa para realizar la respiración celular. O, si el productor es consumido por herbívoros en el siguiente nivel trófico, parte de la energía se transmite a la parte superior de la pirámide. [1] La glucosa almacenada dentro de los productores sirve como alimento para los consumidores , por lo que solo a través de los productores los consumidores pueden acceder a la energía del sol. [1] [7] Algunos ejemplos de productores primarios son las algas , los musgos y otras plantas como las hierbas, los árboles y los arbustos. [1]
Las bacterias quimiosintéticas realizan un proceso similar a la fotosíntesis, pero en lugar de energía del sol utilizan energía almacenada en sustancias químicas como el sulfuro de hidrógeno . [10] [11] Este proceso, conocido como quimiosíntesis , suele ocurrir en las profundidades del océano en respiraderos hidrotermales que producen calor y sustancias químicas como hidrógeno, sulfuro de hidrógeno y metano. [10] Las bacterias quimiosintéticas pueden utilizar la energía de los enlaces del sulfuro de hidrógeno y el oxígeno para convertir el dióxido de carbono en glucosa, liberando agua y azufre en el proceso. [11] Los organismos que consumen las bacterias quimiosintéticas pueden absorber la glucosa y utilizar el oxígeno para realizar la respiración celular, de forma similar a los herbívoros que consumen productores.
Uno de los factores que controla la producción primaria es la cantidad de energía que entra al productor o productores, que se puede medir utilizando la productividad. [12] [13] [1] Solo el uno por ciento de la energía solar entra al productor, el resto rebota o se mueve a través de él. [13] La productividad primaria bruta es la cantidad de energía que el productor realmente obtiene. [13] [14] Generalmente, el 60% de la energía que entra al productor va a la propia respiración del productor. [12] La productividad primaria neta es la cantidad que la planta retiene después de restar la cantidad que utilizó para la respiración celular. [13] Otro factor que controla la producción primaria son los niveles de nutrientes orgánicos/inorgánicos en el agua o el suelo en el que vive el productor. [14]
Producción secundaria
La producción secundaria es el uso de la energía almacenada en las plantas convertida por los consumidores en su propia biomasa. Diferentes ecosistemas tienen diferentes niveles de consumidores, todos terminan con un consumidor superior. La mayor parte de la energía se almacena en la materia orgánica de las plantas, y a medida que los consumidores comen estas plantas absorben esta energía. Esta energía en los herbívoros y omnívoros es luego consumida por los carnívoros . También hay una gran cantidad de energía que está en la producción primaria y termina siendo desperdicio o basura, conocida como detritos. La cadena alimentaria detrítica incluye una gran cantidad de microbios, macroinvertebrados , meiofauna , hongos y bacterias. Estos organismos son consumidos por omnívoros y carnívoros y representan una gran cantidad de producción secundaria. [15] Los consumidores secundarios pueden variar ampliamente en su eficiencia al consumir. [16] Se estima que la eficiencia de la energía que se transmite a los consumidores es de alrededor del 10%. [16] El flujo de energía a través de los consumidores difiere en entornos acuáticos y terrestres.
En ambientes acuáticos
Los heterótrofos contribuyen a la producción secundaria y depende de la productividad primaria y de los productos primarios netos. [16] La producción secundaria es la energía que utilizan los herbívoros y descomponedores y, por lo tanto, depende de la productividad primaria. [16] Principalmente, los herbívoros y descomponedores consumen todo el carbono de dos fuentes orgánicas principales en los ecosistemas acuáticos , autóctonas y alóctonas . [16] El carbono autóctono proviene del interior del ecosistema e incluye plantas acuáticas, algas y fitoplancton. El carbono alóctono de fuera del ecosistema es principalmente materia orgánica muerta del ecosistema terrestre que ingresa al agua. [16] En los ecosistemas fluviales, aproximadamente el 66% de la entrada anual de energía puede lavarse río abajo. La cantidad restante se consume y se pierde en forma de calor. [17]
En ambientes terrestres
La producción secundaria se describe a menudo en términos de niveles tróficos , y si bien esto puede ser útil para explicar las relaciones, enfatiza demasiado las interacciones más raras. Los consumidores a menudo se alimentan en múltiples niveles tróficos. [18] La energía transferida por encima del tercer nivel trófico es relativamente poco importante. [18] La eficiencia de asimilación se puede expresar por la cantidad de alimento que ha comido el consumidor, cuánto asimila el consumidor y qué se expulsa como heces u orina. [19] Mientras que una parte de la energía se utiliza para la respiración, otra parte de la energía se destina a la biomasa en el consumidor. [16] Hay dos cadenas alimentarias principales: la cadena alimentaria primaria es la energía que proviene de los autótrofos y se transmite a los consumidores; y la segunda cadena alimentaria principal es cuando los carnívoros se comen a los herbívoros o descomponedores que consumen la energía autótrofa. [16] Los consumidores se dividen en consumidores primarios, consumidores secundarios y consumidores terciarios. Los carnívoros tienen una asimilación de energía mucho mayor, alrededor del 80%, y los herbívoros tienen una eficiencia mucho menor, de aproximadamente el 20 al 50%. [16] La energía en un sistema puede verse afectada por la emigración/inmigración animal. Los movimientos de los organismos son significativos en los ecosistemas terrestres. [17] El consumo energético de los herbívoros en los ecosistemas terrestres tiene un rango bajo de ~3-7%. [17] El flujo de energía es similar en muchos entornos terrestres. La fluctuación en la cantidad de producto primario neto consumido por los herbívoros es generalmente baja. Esto contrasta enormemente con los entornos acuáticos de lagos y estanques donde los herbívoros tienen un consumo mucho mayor de alrededor del ~33%. [17] Los ectotérmicos y los endotérmicos tienen eficiencias de asimilación muy diferentes. [16]
Detritívoros
Los detritívoros consumen materia orgánica en descomposición y a su vez son consumidos por los carnívoros. [16] La productividad de los depredadores está correlacionada con la productividad de las presas. Esto confirma que la productividad primaria en los ecosistemas afecta a toda la productividad posterior. [20]
Los detritos constituyen una gran parte de la materia orgánica de los ecosistemas. La materia orgánica de los bosques templados está formada principalmente por plantas muertas, aproximadamente el 62 %. [18]
En un ecosistema acuático, la materia de las hojas que cae en los arroyos se moja y comienza a filtrar materia orgánica. Esto sucede con bastante rapidez y atraerá a microbios e invertebrados. Las hojas se pueden descomponer en grandes trozos llamados materia orgánica particulada gruesa (CPOM). [15] La CPOM es colonizada rápidamente por microbios. La meiofauna es extremadamente importante para la producción secundaria en los ecosistemas fluviales. [15] Los microbios que descomponen y colonizan esta materia de las hojas son muy importantes para los detritívoros. Los detritívoros hacen que la materia de las hojas sea más comestible al liberar compuestos de los tejidos; en última instancia, ayuda a ablandarlos. [15] A medida que las hojas se descomponen, el nitrógeno disminuirá ya que la celulosa y la lignina en las hojas son difíciles de descomponer. Por lo tanto, los microbios colonizadores traen nitrógeno para ayudar en la descomposición. La descomposición de las hojas puede depender del contenido inicial de nitrógeno, la estación y la especie de árbol. Las especies de árboles pueden tener variaciones cuando sus hojas caen. Por lo tanto, la descomposición de las hojas ocurre en diferentes momentos, lo que se llama un mosaico de poblaciones microbianas. [15]
El efecto y la diversidad de las especies en un ecosistema se pueden analizar a través de su desempeño y eficiencia. [21] Además, la producción secundaria en los arroyos puede verse muy influenciada por los detritos que caen en ellos; la producción de biomasa y abundancia de fauna bentónica disminuyó un 47-50% adicional durante un estudio de eliminación y exclusión de basura. [20]
Flujo de energía a través de los ecosistemas
Las investigaciones han demostrado que los productores primarios fijan el carbono a tasas similares en todos los ecosistemas. [14] Una vez que el carbono se ha introducido en un sistema como una fuente viable de energía, los mecanismos que gobiernan el flujo de energía a niveles tróficos superiores varían entre los ecosistemas. Entre los ecosistemas acuáticos y terrestres, se han identificado patrones que pueden explicar esta variación y se han dividido en dos vías principales de control: de arriba hacia abajo y de abajo hacia arriba. [22] [23] Los mecanismos que actúan dentro de cada vía regulan en última instancia la estructura de la comunidad y del nivel trófico dentro de un ecosistema en diversos grados. [24] Los controles de abajo hacia arriba implican mecanismos que se basan en la calidad y disponibilidad de los recursos, que controlan la productividad primaria y el flujo posterior de energía y biomasa a niveles tróficos superiores. [23] Los controles de arriba hacia abajo implican mecanismos que se basan en el consumo por parte de los consumidores. [24] [23] Estos mecanismos controlan la tasa de transferencia de energía de un nivel trófico a otro a medida que los herbívoros o depredadores se alimentan de niveles tróficos inferiores. [22]
Ecosistemas acuáticos vs. ecosistemas terrestres
En cada tipo de ecosistema se encuentra una gran variación en el flujo de energía, lo que crea un desafío para identificar la variación entre los tipos de ecosistemas. En un sentido general, el flujo de energía es una función de la productividad primaria con la temperatura, la disponibilidad de agua y la disponibilidad de luz. [25] Por ejemplo, entre los ecosistemas acuáticos, las tasas de producción más altas se encuentran generalmente en los grandes ríos y lagos poco profundos que en los lagos profundos y los arroyos de cabecera claros. [25] Entre los ecosistemas terrestres, las marismas , los pantanos y las selvas tropicales tienen las tasas de producción primaria más altas, mientras que la tundra y los ecosistemas alpinos tienen las más bajas. [25] Las relaciones entre la producción primaria y las condiciones ambientales han ayudado a explicar la variación dentro de los tipos de ecosistemas, lo que permite a los ecólogos demostrar que la energía fluye de manera más eficiente a través de los ecosistemas acuáticos que de los ecosistemas terrestres debido a los diversos controles de abajo hacia arriba y de arriba hacia abajo en juego. [23]
De abajo hacia arriba
La fuerza de los controles de abajo hacia arriba sobre el flujo de energía está determinada por la calidad nutricional, el tamaño y las tasas de crecimiento de los productores primarios en un ecosistema. [14] [22] El material fotosintético es típicamente rico en nitrógeno (N) y fósforo (P) y complementa la alta demanda de N y P de los herbívoros en todos los ecosistemas. [26] La producción primaria acuática está dominada por fitoplancton unicelular pequeño que se compone principalmente de material fotosintético, proporcionando una fuente eficiente de estos nutrientes para los herbívoros. [22] En contraste, las plantas terrestres multicelulares contienen muchas estructuras de celulosa de soporte grandes de alto carbono pero bajo valor nutritivo. [22] Debido a esta diferencia estructural, los productores primarios acuáticos tienen menos biomasa por tejido fotosintético almacenado dentro del ecosistema acuático que en los bosques y pastizales de los ecosistemas terrestres. [22] Esta baja biomasa en relación con el material fotosintético en los ecosistemas acuáticos permite una tasa de rotación más eficiente en comparación con los ecosistemas terrestres. [22] A medida que los herbívoros consumen el fitoplancton, sus mayores tasas de crecimiento y reproducción reemplazan suficientemente la biomasa perdida y, junto con su calidad densa en nutrientes, sustentan una mayor producción secundaria. [22]
Otros factores que afectan la producción primaria incluyen los aportes de N y P, que se producen en mayor magnitud en los ecosistemas acuáticos. [22] Estos nutrientes son importantes para estimular el crecimiento de las plantas y, cuando pasan a niveles tróficos más altos, estimulan la biomasa de los consumidores y la tasa de crecimiento. [23] [25] Si alguno de estos nutrientes escasea, puede limitar la producción primaria general. [15] Dentro de los lagos, el P tiende a ser el nutriente más limitante, mientras que tanto el N como el P limitan la producción primaria en los ríos. [23] Debido a estos efectos limitantes, los aportes de nutrientes pueden aliviar potencialmente las limitaciones en la producción primaria neta de un ecosistema acuático. [24] El material alóctono que se arrastra a un ecosistema acuático introduce N y P, así como energía en forma de moléculas de carbono que son fácilmente absorbidas por los productores primarios. [15] Los mayores aportes y las mayores concentraciones de nutrientes respaldan mayores tasas de producción primaria neta, lo que a su vez respalda una mayor producción secundaria. [26]
De arriba hacia abajo
Los mecanismos de arriba hacia abajo ejercen un mayor control sobre los productores primarios acuáticos debido al papel de los consumidores dentro de una red alimentaria acuática. [24] Entre los consumidores, los herbívoros pueden mediar los impactos de las cascadas tróficas al unir el flujo de energía de los productores primarios a los depredadores en niveles tróficos superiores. [27] En todos los ecosistemas, existe una asociación consistente entre el crecimiento de los herbívoros y la calidad nutricional de los productores. [26] Sin embargo, en los ecosistemas acuáticos, los productores primarios son consumidos por los herbívoros a una tasa cuatro veces mayor que en los ecosistemas terrestres. [22] Aunque este tema es muy debatido, los investigadores han atribuido la distinción en el control de los herbívoros a varias teorías, incluidas las proporciones de tamaño de productor a consumidor y la selectividad de los herbívoros. [7]
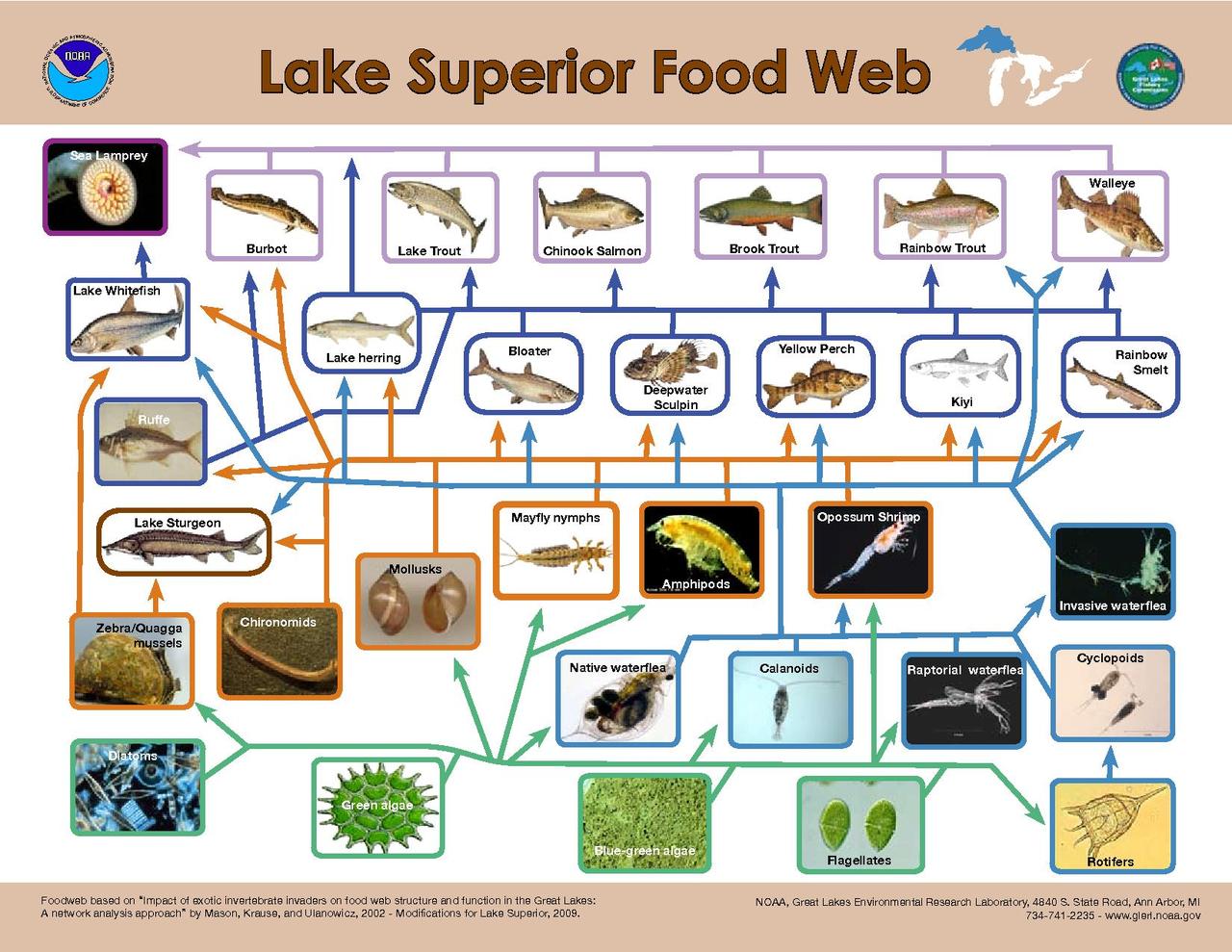
El modelado de controles de arriba hacia abajo sobre los productores primarios sugiere que el mayor control sobre el flujo de energía ocurre cuando la relación de tamaño del consumidor al productor primario es la más alta. [29] La distribución de tamaño de los organismos encontrados dentro de un solo nivel trófico en sistemas acuáticos es mucho más estrecha que la de los sistemas terrestres. [22] En la tierra, el tamaño del consumidor varía desde más pequeño que la planta que consume, como un insecto, hasta significativamente más grande, como un ungulado , mientras que en los sistemas acuáticos, el tamaño corporal del consumidor dentro de un nivel trófico varía mucho menos y está fuertemente correlacionado con la posición trófica. [22] Como resultado, la diferencia de tamaño entre productores y consumidores es consistentemente mayor en ambientes acuáticos que en la tierra, lo que resulta en un control herbívoro más fuerte sobre los productores primarios acuáticos. [22]
Los herbívoros pueden controlar potencialmente el destino de la materia orgánica a medida que se recicla a través de la red alimentaria. [27] Los herbívoros tienden a seleccionar plantas nutritivas mientras evitan las plantas con mecanismos de defensa estructurales . [22] Al igual que las estructuras de soporte, las estructuras de defensa están compuestas de celulosa pobre en nutrientes y alta en carbono. [27] El acceso a fuentes de alimentos nutritivos mejora el metabolismo de los herbívoros y las demandas de energía, lo que lleva a una mayor eliminación de productores primarios. [14] En los ecosistemas acuáticos, el fitoplancton es altamente nutritivo y generalmente carece de mecanismos de defensa. [27] Esto resulta en un mayor control de arriba hacia abajo porque la materia vegetal consumida se libera rápidamente de nuevo en el sistema como desechos orgánicos lábiles . [15] [27] En los ecosistemas terrestres, los productores primarios son menos densos nutricionalmente y es más probable que contengan estructuras de defensa. [22] Debido a que los herbívoros prefieren plantas nutricionalmente densas y evitan plantas o partes de plantas con estructuras de defensa, una mayor cantidad de materia vegetal queda sin consumir dentro del ecosistema. [27] La evitación por parte de los herbívoros de materia vegetal de baja calidad puede ser la razón por la que los sistemas terrestres exhiben un control descendente más débil sobre el flujo de energía. [22]
Véase también
- Red alimentaria : interconexión natural de las cadenas alimentarias
- Estequiometría ecológica
- Energía – Magnitud física
Referencias
- ^ abcdefghijklmn Lindeman RL (1942). "El aspecto trófico-dinámico de la ecología" (PDF) . Ecología . 23 (4): 399–417. Bibcode :1942Ecol...23..399L. doi :10.2307/1930126. JSTOR 1930126. Archivado desde el original (PDF) el 2017-03-29 . Consultado el 2020-12-04 .
- ^ ab Briand F, Cohen JE (noviembre de 1987). "Correlaciones ambientales de la longitud de la cadena alimentaria". Science . 238 (4829). Nueva York, NY: 956–60. Bibcode :1987Sci...238..956B. doi :10.1126/science.3672136. PMID 3672136.
- ^ ab Vander Zanden MJ, Shuter BJ, Lester N, Rasmussen JB (octubre de 1999). "Patrones de longitud de la cadena alimentaria en lagos: un estudio de isótopos estables". The American Naturalist . 154 (4): 406–416. doi :10.1086/303250. PMID 10523487. S2CID 4424697.
- ^ Sharma JP (2009). Estudios ambientales (3.ª ed.). Nueva Delhi: University Science Press. ISBN 978-81-318-0641-8.OCLC 908431622 .
- ^ Van Ness HC (1969). Entendiendo la termodinámica (ed. Dover). Nueva York: Dover Publications, Inc. ISBN 978-1-62198-625-6.OCLC 849744641 .
- ^ "Ciclo del carbono". Archivado desde el original el 12 de agosto de 2006.
- ^ abcd Whitmarsh J, Govindjee (1999). "El proceso fotosintético". En Singhal GS, Renger G, Sopory SK, Irrgang KD, Govindjee (eds.). Conceptos de fotobiología: fotosíntesis y fotomorfogénesis . Boston: Kluwer Academic Publishers . págs. 11–51. ISBN 978-0-7923-5519-9..
100×10 15 gramos de carbono/año fijados por organismos fotosintéticos, lo que equivale a 4×10 18 kJ/año = 4×10 21 J/año de energía libre almacenada como carbono reducido.
- ^ Teal JM (1962). "Flujo de energía en el ecosistema de marismas de Georgia". Ecología . 43 (4): 614–624. Bibcode :1962Ecol...43..614T. doi :10.2307/1933451. JSTOR 1933451.
- ^ Morris J, Hartl DL, Knoll AH, Lue R, Michael M (2019). Biología: cómo funciona la vida (3.ª ed.). WH Freeman. ISBN 978-1-319-01763-7.
- ^ ab Kellermann MY, Wegener G, Elvert M, Yoshinaga MY, Lin YS, Holler T, et al. (noviembre de 2012). "Autotrofia como modo predominante de fijación de carbono en comunidades microbianas anaeróbicas oxidantes de metano". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 109 (47): 19321–6. Bibcode :2012PNAS..10919321K. doi : 10.1073/pnas.1208795109 . PMC 3511159 . PMID 23129626.
- ^ ab Cavanaugh CM, Gardiner SL, Jones ML, Jannasch HW, Waterbury JB (julio de 1981). "Células procariotas en el gusano tubular de los respiraderos hidrotermales Riftia pachyptila Jones: posibles simbiontes quimioautotróficos". Science . 213 (4505). Nueva York, NY: 340–2. Bibcode :1981Sci...213..340C. doi :10.1126/science.213.4505.340. PMID 17819907.
- ^ ab Amthor JS, Baldocchi DD (2001). "Respiración de plantas superiores terrestres y producción primaria neta". Productividad global terrestre : 33–59. doi :10.1016/B978-012505290-0/50004-1. ISBN 978-0-12-505290-0.
- ^ abcd Sigman DM, Hain MP (2012). "La productividad biológica del océano" (PDF) . Nature Education Knowledge . 3 (6): 1–6.[ enlace muerto permanente ]
- ^ abcde Cebrian J (octubre de 1999). "Patrones en el destino de la producción en comunidades vegetales". The American Naturalist . 154 (4): 449–468. doi :10.1086/303244. PMID 10523491. S2CID 4384243.
- ^ abcdefgh Allan JD, Castillo MM (2007). Ecología de los cursos de agua: estructura y función de las aguas corrientes (2.ª ed.). Dordrecht: Springer. ISBN 978-1-4020-5582-9.OCLC 144222191 .
- ^ abcdefghijk Smith TM, Smith RL (2015). Elementos de ecología (novena edición). Boston: Pearson. ISBN 978-1-292-07741-3.OCLC 914328590 .
- ^ abcd Fisher SG, Likens GE (febrero de 1973). "Flujo de energía en Bear Brook, New Hampshire: un enfoque integrador del metabolismo de los ecosistemas fluviales". Monografías ecológicas . 43 (4): 421–439. Bibcode :1973EcoM...43..421F. doi :10.2307/1942301. JSTOR 1942301.
- ^ abc Hairston Jr NG, Hairston Sr NG (septiembre de 1993). "Relaciones de causa y efecto en el flujo de energía, la estructura trófica y las interacciones interespecíficas". The American Naturalist . 142 (3): 379–411. doi :10.1086/285546. hdl : 1813/57238 . S2CID 55279332.
- ^ Sanders D, Moser A, Newton J, van Veen FJ (16 de marzo de 2016). "La eficiencia de asimilación trófica aumenta notablemente en niveles tróficos superiores en la cadena alimentaria de cuatro niveles huésped-parasitoide". Actas de la Royal Society B: Ciencias Biológicas . 283 (1826): 20153043. doi :10.1098/rspb.2015.3043. ISSN 0962-8452. PMC 4810866 . PMID 26962141.
- ^ ab Wallace JB, Eggert SL, Meyer JL, Webster JR (noviembre de 1999). "Efectos de la limitación de recursos en un ecosistema basado en detríticos". Monografías ecológicas . 69 (4): 409–42. doi :10.1890/0012-9615(1999)069[0409:eorloa]2.0.co;2.
- ^ Cardinale BJ, Srivastava DS, Duffy JE, Wright JP, Downing AL, Sankaran M, et al. (octubre de 2006). "Efectos de la biodiversidad en el funcionamiento de los grupos tróficos y los ecosistemas". Nature . 443 (7114): 989–92. Bibcode :2006Natur.443..989C. doi :10.1038/nature05202. PMID 17066035. S2CID 4426751.
- ^ abcdefghijklmnop Shurin JB, Gruner DS, Hillebrand H (enero de 2006). "¿Todo mojado o seco? Diferencias reales entre las redes alimentarias acuáticas y terrestres". Actas. Ciencias biológicas . 273 (1582): 1–9. doi :10.1098/rspb.2005.3377. PMC 1560001. PMID 16519227 .
- ^ abcdef La Pierre K, Hanley T (2015). Ecología trófica: interacciones de abajo hacia arriba y de arriba hacia abajo en los sistemas acuáticos y terrestres . Cambridge University Press. págs. 55–85. ISBN 978-1-316-29969-2.
- ^ abcd Gruner DS, Smith JE, Seabloom EW, Sandin SA, Ngai JT, Hillebrand H, et al. (julio de 2008). "Una síntesis intersistémica del control de los recursos de nutrientes y consumidores sobre la biomasa del productor". Ecology Letters . 11 (7): 740–55. Bibcode :2008EcolL..11..740G. doi :10.1111/j.1461-0248.2008.01192.x. PMID 18445030.
- ^ abcd Ricklefs RE, Miller GL (2000). Ecología (4.ª ed.). Nueva York: WH Freeman & Co. ISBN 0-7167-2829-X.OCLC 40734932 .
- ^ abc Cebrian J, Lartigue J (2004). "Patrones de herbivoría y descomposición en ecosistemas acuáticos y terrestres". Monografías ecológicas . 74 (2): 237–259. Código Bibliográfico :2004EcoM...74..237C. doi :10.1890/03-4019.
- ^ abcdef Schmitz OJ (diciembre de 2008). "Herbivoría desde individuos hasta ecosistemas". Revista anual de ecología, evolución y sistemática . 39 (1): 133–152. doi :10.1146/annurev.ecolsys.39.110707.173418.
- ^ Krause AE, Frank KA, Mason DM, Ulanowicz RE, Taylor WW (noviembre de 2003). "Compartimentos revelados en la estructura de la red alimentaria". Nature . 426 (6964): 282–5. Bibcode :2003Natur.426..282K. doi :10.1038/nature02115. hdl : 2027.42/62960 . PMID 14628050. S2CID 1752696.
- ^ Shurin JB, Seabloom EW (2005). "La fuerza de las cascadas tróficas en los ecosistemas: predicciones a partir de la alometría y la energética". Journal of Animal Ecology . 74 (6): 1029–1038. Bibcode :2005JAnEc..74.1029S. doi : 10.1111/j.1365-2656.2005.00999.x . ISSN 1365-2656.
Lectura adicional
- Podolinsky S (2004). "Socialismo y la unidad de las fuerzas físicas". Organización y medio ambiente . 17 (1): 61–75. doi :10.1177/1086026603262092. S2CID 144332347.
- Weiner DR (2000). Modelos de la naturaleza: ecología, conservación y revolución cultural en la Rusia soviética . EE.UU.: University of Pittsburgh Press .
