Cicatrización de heridas
| Abrasión de la mano | ||||
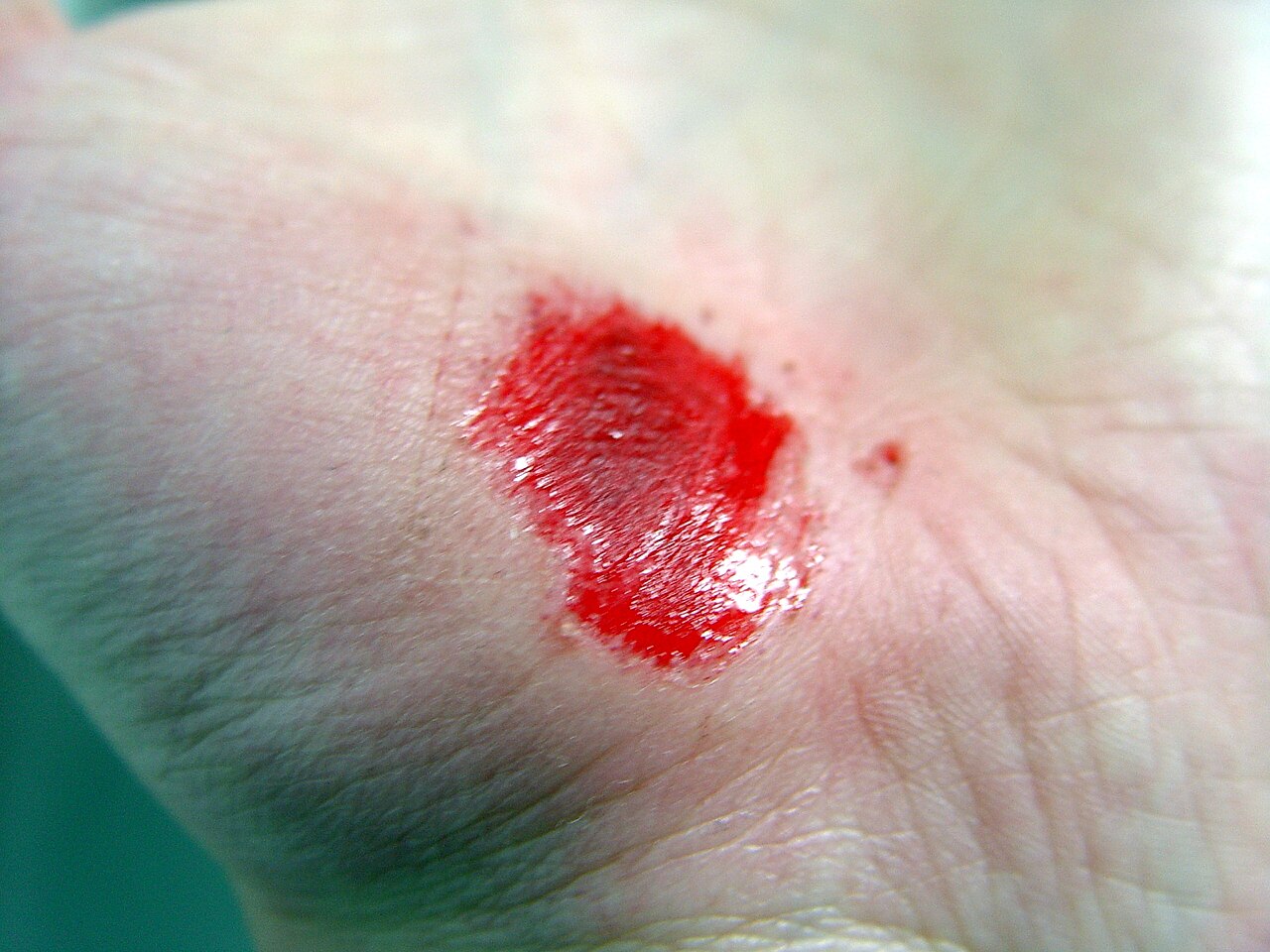 |  |  |  | |
| Lesión inicial | 3 días | 17 días | 30 días | |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
La cicatrización de heridas se refiere al reemplazo de tejido destruido o dañado por tejido nuevo producido por un organismo vivo. [1]
En la piel no dañada, la epidermis ( capa epitelial superficial ) y la dermis ( capa conectiva más profunda ) forman una barrera protectora contra el entorno externo. Cuando se rompe la barrera, se pone en marcha una secuencia regulada de eventos bioquímicos para reparar el daño. [1] [2] Este proceso se divide en fases predecibles: coagulación sanguínea ( hemostasia ), inflamación , crecimiento tisular ( proliferación celular ) y remodelación tisular (maduración y diferenciación celular ). La coagulación sanguínea puede considerarse parte de la etapa de inflamación en lugar de una etapa separada. [3]
{kind=link}

El proceso de cicatrización de heridas no sólo es complejo sino también frágil y susceptible de interrupción o falla que conduzca a la formación de heridas crónicas que no cicatrizan . Los factores que contribuyen a la falta de cicatrización de heridas crónicas son la diabetes, las enfermedades venosas o arteriales, las infecciones y las deficiencias metabólicas de la vejez. [4]
El cuidado de las heridas estimula y acelera su curación mediante la limpieza y la protección contra nuevas lesiones o infecciones. Según las necesidades de cada paciente, puede abarcar desde los primeros auxilios más sencillos hasta especialidades de enfermería completas, como enfermería de heridas, ostomía y continencia y atención en centros de quemados .
Etapas
- Hemostasia (coagulación de la sangre): en los primeros minutos de la lesión, las plaquetas de la sangre comienzan a adherirse al lugar lesionado. Adoptan una forma amorfa, más adecuada para la coagulación, y liberan señales químicas para promover la coagulación. Esto da como resultado la activación de la fibrina , que forma una malla y actúa como "pegamento" para unir las plaquetas entre sí. Esto produce un coágulo que sirve para tapar la rotura del vaso sanguíneo, lo que ralentiza o previene un mayor sangrado. [5] [6]
- Inflamación: Durante esta fase, se eliminan las células dañadas y muertas, junto con las bacterias y otros patógenos o desechos. Esto sucede a través del proceso de fagocitosis , donde los glóbulos blancos engullen los desechos y los destruyen. Los factores de crecimiento derivados de las plaquetas se liberan en la herida y provocan la migración y división de las células durante la fase proliferativa.
- Proliferación (crecimiento de tejido nuevo): En esta fase, se produce la angiogénesis , la deposición de colágeno , la formación de tejido de granulación , la epitelización y la contracción de la herida. [7] En la angiogénesis, las células endoteliales vasculares forman nuevos vasos sanguíneos. [8] En la fibroplasia y la formación de tejido de granulación, los fibroblastos crecen y forman una nueva matriz extracelular (ECM) provisional mediante la excreción de colágeno y fibronectina . [7] Al mismo tiempo, se produce la reepitelización de la epidermis, en la que las células epiteliales proliferan y "se arrastran" sobre el lecho de la herida, proporcionando cobertura para el nuevo tejido. [9] En la contracción de la herida, los miofibroblastos disminuyen el tamaño de la herida agarrando los bordes de la herida y contrayéndose utilizando un mecanismo que se asemeja al de las células del músculo liso. Cuando las funciones de las células están cerca de completarse, las células innecesarias experimentan apoptosis . [7]
- Maduración (remodelación): Durante la maduración y la remodelación, el colágeno se realinea a lo largo de líneas de tensión y las células que ya no son necesarias se eliminan mediante muerte celular programada o apoptosis .
{kind=link}

Sincronización y reepitelización
El momento de la curación de las heridas es importante. El momento de la reepitelización de la herida puede determinar el resultado de la curación. [11] Si la epitelización del tejido sobre una zona denudada es lenta, se formará una cicatriz en el transcurso de muchas semanas o meses; [12] [13] Si la epitelización de una zona herida es rápida, la curación dará lugar a la regeneración. [13]
Fase temprana vs fase celular
{kind=link}

La cicatrización de heridas se divide clásicamente en hemostasia , inflamación, proliferación y remodelación. Aunque es un modelo útil, emplea una superposición considerable entre las fases individuales. Recientemente se ha descrito un modelo complementario [1] en el que se delinean con mayor claridad los numerosos elementos de la cicatrización de heridas. La importancia de este nuevo modelo se hace más evidente a través de su utilidad en los campos de la medicina regenerativa y la ingeniería de tejidos (consulte la sección Investigación y desarrollo a continuación). En este modelo, el proceso de cicatrización de heridas se divide en dos fases principales: la fase temprana y la fase celular : [1]
La fase temprana, que comienza inmediatamente después de una lesión en la piel, involucra eventos moleculares y celulares en cascada que conducen a la hemostasia y la formación de una matriz extracelular temprana e improvisada que proporciona una etapa estructural para la adhesión celular y la proliferación celular posterior.
La fase celular involucra varios tipos de células que trabajan juntas para generar una respuesta inflamatoria, sintetizar tejido de granulación y restaurar la capa epitelial. [1] Las subdivisiones de la fase celular son:
- Macrófagos y componentes inflamatorios (en 1-2 días)
- Interacción epitelial - mesenquimal : reepitelización ( cambio de fenotipo en cuestión de horas, la migración comienza el día 1 o 2)
- Fibroblastos y miofibroblastos : alineación progresiva, producción de colágeno y contracción de la matriz (entre el día 4 y el día 14)
- Células endoteliales y angiogénesis (comienza el día 4)
- Matriz dérmica: elementos de fabricación (comienza el día 4 y dura 2 semanas) y alteración/remodelación (comienza después de la semana 2 y dura semanas o meses, según el tamaño de la herida). [1]
Fase inflamatoria
Justo antes de que se inicie la fase inflamatoria, se produce la cascada de coagulación para lograr la hemostasia , o la detención de la pérdida de sangre mediante un coágulo de fibrina. A continuación, se liberan diversos factores solubles (entre ellos, quimiocinas y citocinas) para atraer células que fagociten restos, bacterias y tejido dañado, además de liberar moléculas señalizadoras que inician la fase proliferativa de la cicatrización de las heridas.
Cascada de coagulación
Cuando el tejido se hiere por primera vez, la sangre entra en contacto con el colágeno , lo que hace que las plaquetas sanguíneas comiencen a secretar factores inflamatorios. [15] Las plaquetas también expresan glicoproteínas pegajosas en sus membranas celulares que les permiten agregarse y formar una masa. [7]
La fibrina y la fibronectina se unen entre sí y forman un tapón que atrapa proteínas y partículas y evita una mayor pérdida de sangre. [16] Este tapón de fibrina-fibronectina también es el principal soporte estructural de la herida hasta que se deposita el colágeno. [7] Las células migratorias utilizan este tapón como matriz para arrastrarse, y las plaquetas se adhieren a él y secretan factores. [7] El coágulo finalmente se lisa y se reemplaza con tejido de granulación y luego con colágeno.
Las plaquetas, las células presentes en mayor número poco después de que se produce una herida, liberan mediadores en la sangre, incluidas las citocinas y los factores de crecimiento . [15] Los factores de crecimiento estimulan a las células para que aceleren su tasa de división. Las plaquetas liberan otros factores proinflamatorios como la serotonina , la bradicinina , las prostaglandinas , las prostaciclinas , el tromboxano y la histamina , [3] que cumplen varias funciones, incluido el aumento de la proliferación y la migración celular al área y la dilatación y la porosidad de los vasos sanguíneos . En muchos sentidos, las plaquetas extravasadas en el traumatismo realizan una función similar a los macrófagos tisulares y los mastocitos expuestos a las firmas moleculares microbianas en la infección: se activan y secretan mediadores moleculares (aminas vasoactivas, eicosanoides y citocinas ) que inician el proceso inflamatorio .
Vasoconstricción y vasodilatación
Inmediatamente después de que se rompe un vaso sanguíneo , las membranas celulares rotas liberan factores inflamatorios como tromboxanos y prostaglandinas que hacen que el vaso se espasme para evitar la pérdida de sangre y para acumular células y factores inflamatorios en el área. [3] Esta vasoconstricción dura de cinco a diez minutos y es seguida por vasodilatación , un ensanchamiento de los vasos sanguíneos, que alcanza su punto máximo aproximadamente a los 20 minutos después de la herida. [3] La vasodilatación es el resultado de factores liberados por las plaquetas y otras células. El principal factor involucrado en causar vasodilatación es la histamina . [3] [15] La histamina también hace que los vasos sanguíneos se vuelvan porosos, lo que permite que el tejido se vuelva edematoso porque las proteínas del torrente sanguíneo se filtran al espacio extravascular, lo que aumenta su carga osmolar y atrae agua hacia el área. [3] El aumento de la porosidad de los vasos sanguíneos también facilita la entrada de células inflamatorias como los leucocitos en el sitio de la herida desde el torrente sanguíneo . [17] [18]
Neutrófilos polimorfonucleares
En el plazo de una hora tras la herida, los neutrófilos polimorfonucleares (PMN) llegan al lugar de la herida y se convierten en las células predominantes en la herida durante los dos primeros días posteriores a la lesión, con números especialmente altos en el segundo día. [19] Son atraídos al sitio por la fibronectina, los factores de crecimiento y sustancias como las cininas . Los neutrófilos fagocitan los desechos y matan las bacterias liberando radicales libres en lo que se denomina una explosión respiratoria . [20] [21] También limpian la herida al secretar proteasas que descomponen el tejido dañado. Los neutrófilos funcionales en el sitio de la herida solo tienen una vida útil de alrededor de dos días, por lo que generalmente experimentan apoptosis una vez que han completado sus tareas y son engullidos y degradados por los macrófagos . [22]
Otros leucocitos que ingresan al área incluyen las células T auxiliares , que secretan citocinas para hacer que más células T se dividan y para aumentar la inflamación y mejorar la vasodilatación y la permeabilidad de los vasos. [17] [23] Las células T también aumentan la actividad de los macrófagos. [17]
Macrófagos
Una de las funciones de los macrófagos es fagocitar otros fagocitos gastados , [24] bacterias y tejido dañado, [19] y también desbridan el tejido dañado liberando proteasas. [25]
Los macrófagos funcionan en la regeneración [26] [27] y son esenciales para la cicatrización de heridas. [19] Son estimulados por el bajo contenido de oxígeno de su entorno para producir factores que inducen y aceleran la angiogénesis [20] y también estimulan las células que reepitelizan la herida, crean tejido de granulación y depositan una nueva matriz extracelular . [28] Al secretar estos factores, los macrófagos contribuyen a impulsar el proceso de cicatrización de heridas a la siguiente fase. Reemplazan a los PMN como células predominantes en la herida dos días después de la lesión. [24]
El bazo contiene la mitad de los monocitos del cuerpo en reserva listos para ser desplegados en el tejido lesionado. [29] [30] Atraídos al sitio de la herida por los factores de crecimiento liberados por las plaquetas y otras células, los monocitos del torrente sanguíneo ingresan al área a través de las paredes de los vasos sanguíneos. [31] La cantidad de monocitos en la herida alcanza su punto máximo entre un día y un día y medio después de que se produce la lesión. [23] Una vez que están en el sitio de la herida, los monocitos maduran y se convierten en macrófagos. Los macrófagos también secretan una serie de factores, como factores de crecimiento y otras citocinas, especialmente durante el tercer y cuarto día posteriores a la herida. Estos factores atraen a la zona a las células involucradas en la etapa de proliferación de la curación. [15]
En la cicatrización de heridas que resultan en una reparación incompleta, se produce una contracción de la cicatriz, lo que produce diferentes grados de imperfecciones estructurales, deformidades y problemas de flexibilidad. [32] Los macrófagos pueden restringir la fase de contracción. [27] Los científicos han informado que la eliminación de los macrófagos de una salamandra resultó en el fracaso de una respuesta de regeneración típica (regeneración de las extremidades), provocando en cambio una respuesta de reparación (cicatrización). [33] [34]
Disminución de la fase inflamatoria
A medida que la inflamación disminuye, se secretan menos factores inflamatorios, los existentes se descomponen y se reduce la cantidad de neutrófilos y macrófagos en el sitio de la herida. [19] Estos cambios indican que la fase inflamatoria está terminando y la fase proliferativa está en marcha. [19] La evidencia in vitro, obtenida utilizando el modelo equivalente dérmico , sugiere que la presencia de macrófagos en realidad retrasa la contracción de la herida y, por lo tanto, la desaparición de los macrófagos de la herida puede ser esencial para que ocurran las fases posteriores. [27]
Debido a que la inflamación desempeña un papel en la lucha contra las infecciones, la limpieza de los residuos y la inducción de la fase de proliferación, es una parte necesaria de la curación. Sin embargo, la inflamación puede provocar daños en los tejidos si dura demasiado tiempo. [7] Por lo tanto, la reducción de la inflamación es con frecuencia un objetivo en los entornos terapéuticos. La inflamación dura mientras haya residuos en la herida. Por lo tanto, si el sistema inmunológico del individuo está comprometido y no puede eliminar los residuos de la herida y/o si hay exceso de detritos, tejido desvitalizado o biopelícula microbiana en la herida, estos factores pueden causar una fase inflamatoria prolongada e impedir que la herida comience adecuadamente la fase de proliferación de la curación. Esto puede dar lugar a una herida crónica .
Fase proliferativa
Aproximadamente dos o tres días después de que se produce la herida, los fibroblastos comienzan a ingresar al sitio de la herida, lo que marca el inicio de la fase proliferativa incluso antes de que haya terminado la fase inflamatoria. [35] Al igual que en las otras fases de la cicatrización de heridas, los pasos de la fase proliferativa no ocurren en serie, sino que se superponen parcialmente en el tiempo.
Angiogénesis
También llamado neovascularización, el proceso de angiogénesis ocurre simultáneamente con la proliferación de fibroblastos cuando las células endoteliales migran al área de la herida. [36] Debido a que la actividad de los fibroblastos y las células epiteliales requiere oxígeno y nutrientes, la angiogénesis es imperativa para otras etapas en la cicatrización de heridas, como la migración epidérmica y de fibroblastos. El tejido en el que se ha producido la angiogénesis normalmente se ve rojo (es eritematoso ) debido a la presencia de capilares . [36]
La angiogénesis ocurre en fases superpuestas en respuesta a la inflamación:
- Periodo de latencia: Durante la fase hemostática e inflamatoria del proceso de cicatrización de la herida, la vasodilatación y la permeabilización permiten la extravasación de leucocitos y el desbridamiento fagocítico y la descontaminación de la zona de la herida. La hinchazón del tejido favorece la angiogénesis posterior al expandir y aflojar la matriz extracelular de colágeno existente.
- Activación endotelial : a medida que los macrófagos de la herida pasan del modo inflamatorio al modo de curación, comienzan a secretar factores quimiotácticos y de crecimiento endoteliales para atraer a las células endoteliales adyacentes. Las células endoteliales activadas responden retrayendo y reduciendo las uniones celulares, desprendiéndose del endotelio en el que se encuentran. Es característico que las células endoteliales activadas presenten nucléolos agrandados.
- Degradación de la membrana basal endotelial : los macrófagos de la herida, los mastocitos y las propias células endoteliales secretan proteasas para descomponer la lámina basal vascular existente.
- Brote vascular : con la ruptura de la membrana basal endotelial, las células endoteliales desprendidas de los capilares preexistentes y las vénulas poscapilares pueden dividirse y migrar quimiotácticamente hacia la herida, formando nuevos vasos en el proceso. El brote vascular puede verse favorecido por la hipoxia ambiental y la acidosis en el entorno de la herida, ya que la hipoxia estimula el factor de transcripción endotelial, factor inducible por hipoxia (HIF), para transactivar genes angiogénicos como VEGF y GLUT1. Los vasos brotados pueden autoorganizarse en morfologías luminales, y la fusión de canales ciegos da lugar a nuevas redes capilares.
- Maduración vascular : el endotelio de los vasos madura depositando nueva matriz extracelular endotelial, seguida de la formación de la lámina basal. Por último, el vaso establece una capa de pericitos.
Las células madre de las células endoteliales , que se originan a partir de partes de vasos sanguíneos no lesionados, desarrollan pseudópodos y avanzan a través de la matriz extracelular hacia el sitio de la herida para establecer nuevos vasos sanguíneos. [20]
Las células endoteliales son atraídas al área de la herida por la fibronectina que se encuentra en la costra de fibrina y quimiotácticamente por factores angiogénicos liberados por otras células, [37] por ejemplo, de los macrófagos y plaquetas cuando se encuentran en un entorno con poco oxígeno. El crecimiento y la proliferación endotelial también se estimulan directamente por la hipoxia y la presencia de ácido láctico en la herida. [35] Por ejemplo, la hipoxia estimula el factor de transcripción endotelial , factor inducible por hipoxia (HIF) para transactivar un conjunto de genes proliferativos que incluyen el factor de crecimiento endotelial vascular (VEGF) y el transportador de glucosa 1 (GLUT1).
Para migrar, las células endoteliales necesitan colagenasas y activador del plasminógeno para degradar el coágulo y parte de la matriz extracelular. [3] [19] Las metaloproteinasas dependientes del zinc digieren la membrana basal y la matriz extracelular para permitir la migración celular, la proliferación y la angiogénesis. [38]
Cuando los macrófagos y otras células productoras de factores de crecimiento ya no se encuentran en un ambiente hipóxico lleno de ácido láctico, dejan de producir factores angiogénicos. [20] Por lo tanto, cuando el tejido está adecuadamente perfundido , la migración y proliferación de células endoteliales se reduce. Finalmente, los vasos sanguíneos que ya no son necesarios mueren por apoptosis . [37]
Fibroplasia y formación de tejido de granulación
Simultáneamente con la angiogénesis, los fibroblastos comienzan a acumularse en el lugar de la herida. Los fibroblastos comienzan a ingresar al lugar de la herida entre dos y cinco días después de la herida, cuando la fase inflamatoria está terminando, y su número alcanza su punto máximo entre una y dos semanas después de la herida. [19] Al final de la primera semana, los fibroblastos son las células principales de la herida. [3] La fibroplasia termina entre dos y cuatro semanas después de la herida.
Como modelo, el mecanismo de la fibroplasia puede conceptualizarse como un proceso análogo a la angiogénesis (ver arriba), sólo que el tipo de célula involucrada son fibroblastos en lugar de células endoteliales. Inicialmente hay una fase latente donde la herida sufre exudación plasmática, descontaminación inflamatoria y desbridamiento. El edema aumenta la accesibilidad histológica de la herida para la posterior migración fibroplástica. En segundo lugar, a medida que la inflamación se acerca a su finalización, los macrófagos y los mastocitos liberan factores de crecimiento de fibroblastos y quimiotácticos para activar los fibroblastos del tejido adyacente. Los fibroblastos en esta etapa se desprenden de las células circundantes y de la matriz extracelular. Los fagocitos liberan además proteasas que descomponen la matriz extracelular del tejido vecino, liberando a los fibroblastos activados para que proliferen y migren hacia la herida. La diferencia entre la brotación vascular y la proliferación de fibroblastos es que la primera se mejora con la hipoxia, mientras que la segunda se inhibe con la hipoxia. El tejido conectivo fibroblástico depositado madura secretando matriz extracelular en el espacio extracelular, formando tejido de granulación (ver abajo). Por último, el colágeno se deposita en la matriz extracelular (ECM).
En los primeros dos o tres días después de la lesión, los fibroblastos migran y proliferan principalmente, mientras que más tarde, son las células principales que depositan la matriz de colágeno en el sitio de la herida. [3] Se cree que los orígenes de estos fibroblastos son del tejido cutáneo adyacente no lesionado (aunque nueva evidencia sugiere que algunos se derivan de células madre/precursores adultos circulantes transmitidos por la sangre). [39] Inicialmente, los fibroblastos utilizan las fibras de reticulación de fibrina (bien formadas al final de la fase inflamatoria) para migrar a través de la herida, adhiriéndose posteriormente a la fibronectina. [37] Luego, los fibroblastos depositan sustancia fundamental en el lecho de la herida y, más tarde, colágeno, al que pueden adherirse para migrar. [15]
El tejido de granulación funciona como tejido rudimentario y comienza a aparecer en la herida ya durante la fase inflamatoria, de dos a cinco días después de la herida, y continúa creciendo hasta que se cubre el lecho de la herida. El tejido de granulación consiste en nuevos vasos sanguíneos, fibroblastos, células inflamatorias, células endoteliales, miofibroblastos y los componentes de una nueva matriz extracelular provisional (ECM). La ECM provisional es diferente en composición de la ECM en el tejido normal y sus componentes se originan a partir de fibroblastos. [28] Dichos componentes incluyen fibronectina, colágeno, glicosaminoglicanos , elastina , glicoproteínas y proteoglicanos . [37] Sus componentes principales son la fibronectina y el hialuronano , que crean una matriz muy hidratada y facilitan la migración celular. [31] Posteriormente, esta matriz provisional se reemplaza con una ECM que se parece más a la que se encuentra en el tejido no lesionado.
Los factores de crecimiento ( PDGF , TGF-β ) y la fibronectina estimulan la proliferación, la migración al lecho de la herida y la producción de moléculas de matriz extracelular por parte de los fibroblastos. Los fibroblastos también secretan factores de crecimiento que atraen a las células epiteliales al sitio de la herida. La hipoxia también contribuye a la proliferación de fibroblastos y a la excreción de factores de crecimiento, aunque una cantidad insuficiente de oxígeno inhibirá su crecimiento y la deposición de componentes de la matriz extracelular, y puede provocar una cicatrización fibrótica excesiva .
Deposición de colágeno
Una de las funciones más importantes de los fibroblastos es la producción de colágeno . [36]
La deposición de colágeno es importante porque aumenta la resistencia de la herida; antes de que se deposite, lo único que mantiene la herida cerrada es el coágulo de fibrina-fibronectina, que no proporciona mucha resistencia a las lesiones traumáticas . [20] Además, las células implicadas en la inflamación, la angiogénesis y la construcción del tejido conectivo se adhieren, crecen y se diferencian en la matriz de colágeno depositada por los fibroblastos. [40]
El colágeno tipo III y la fibronectina generalmente comienzan a producirse en cantidades apreciables en algún momento entre aproximadamente 10 horas [41] y 3 días, [37] dependiendo principalmente del tamaño de la herida. Su deposición alcanza su punto máximo entre una y tres semanas. [28] Son las sustancias tensoras predominantes hasta la fase posterior de maduración, en la que son reemplazadas por el colágeno tipo I, más fuerte .
Mientras los fibroblastos producen colágeno nuevo, las colagenasas y otros factores lo degradan. Poco después de la herida, la síntesis supera a la degradación, por lo que los niveles de colágeno en la herida aumentan, pero más tarde la producción y la degradación se igualan, por lo que no hay una ganancia neta de colágeno. [20] Esta homeostasis señala el inicio de la fase de maduración posterior. La granulación cesa gradualmente y los fibroblastos disminuyen en número en la herida una vez que su trabajo ha terminado. [42] Al final de la fase de granulación, los fibroblastos comienzan a cometer apoptosis, convirtiendo el tejido de granulación de un entorno rico en células a uno que consiste principalmente en colágeno. [3]
Epitelización
La formación de tejido de granulación en una herida abierta permite que se produzca la fase de reepitelización, ya que las células epiteliales migran a través del nuevo tejido para formar una barrera entre la herida y el medio ambiente. [37] Los queratinocitos basales de los bordes de la herida y los apéndices dérmicos , como los folículos pilosos , las glándulas sudoríparas y las glándulas sebáceas (aceite) son las principales células responsables de la fase de epitelización de la cicatrización de la herida. [42] Avanzan en una lámina a través del sitio de la herida y proliferan en sus bordes, cesando el movimiento cuando se encuentran en el medio. En la curación que da lugar a una cicatriz, las glándulas sudoríparas, los folículos pilosos [43] [44] y los nervios no se forman. Con la falta de folículos pilosos, nervios y glándulas sudoríparas, la herida y la cicatriz que se está curando suponen un desafío para el cuerpo en lo que respecta al control de la temperatura. [44]
Los queratinocitos migran sin proliferar primero. [45] La migración puede comenzar tan pronto como unas horas después de la herida. Sin embargo, las células epiteliales necesitan tejido viable para migrar, por lo que si la herida es profunda, primero debe llenarse con tejido de granulación. [46] Por lo tanto, el momento del inicio de la migración es variable y puede ocurrir aproximadamente un día después de la herida. [47] Las células en los márgenes de la herida proliferan en el segundo y tercer día después de la herida para proporcionar más células para la migración. [28]
Si no se rompe la membrana basal , las células epiteliales se reemplazan en tres días por división y migración ascendente de células en el estrato basal de la misma manera que ocurre en la piel no lesionada. [37] Sin embargo, si la membrana basal está arruinada en el sitio de la herida, la reepitelización debe ocurrir desde los márgenes de la herida y desde los apéndices de la piel, como los folículos pilosos y las glándulas sudoríparas y sebáceas que ingresan a la dermis que están revestidas con queratinocitos viables. [28] Si la herida es muy profunda, los apéndices de la piel también pueden arruinarse y la migración solo puede ocurrir desde los bordes de la herida. [46]
La migración de los queratinocitos sobre el sitio de la herida es estimulada por la falta de inhibición de contacto y por sustancias químicas como el óxido nítrico . [48] Antes de que comiencen a migrar, las células deben disolver sus desmosomas y hemidesmosomas , que normalmente anclan las células mediante filamentos intermedios en su citoesqueleto a otras células y a la matriz extracelular. [23] Las proteínas receptoras transmembrana llamadas integrinas , que están hechas de glicoproteínas y normalmente anclan la célula a la membrana basal mediante su citoesqueleto , se liberan de los filamentos intermedios de la célula y se reubican en filamentos de actina para servir como accesorios a la matriz extracelular para los pseudópodos durante la migración. [23] De este modo, los queratinocitos se desprenden de la membrana basal y pueden entrar en el lecho de la herida. [35]
Antes de comenzar a migrar, los queratinocitos cambian de forma, volviéndose más largos y planos y extendiendo procesos celulares como lamelipodios y procesos anchos que parecen volantes. [31] Se forman filamentos de actina y pseudópodos . [35] Durante la migración, las integrinas en el pseudópodo se unen a la matriz extracelular, y los filamentos de actina en la proyección tiran de la célula. [23] La interacción con moléculas en la matriz extracelular a través de las integrinas promueve aún más la formación de filamentos de actina, lamelipodios y filopodios . [23]
Las células epiteliales trepan unas sobre otras para migrar. [42] Esta capa creciente de células epiteliales a menudo se denomina lengua epitelial. [45] Las primeras células que se adhieren a la membrana basal forman el estrato basal . Estas células basales continúan migrando a través del lecho de la herida y las células epiteliales que se encuentran por encima de ellas también se deslizan. [45] Cuanto más rápidamente se produzca esta migración, menos cicatriz habrá. [49]
La fibrina , el colágeno y la fibronectina en la matriz extracelular pueden enviar señales adicionales a las células para que se dividan y migren. Al igual que los fibroblastos, los queratinocitos migratorios utilizan la fibronectina reticulada con la fibrina que se depositó en la inflamación como un sitio de unión para atravesarla. [25] [31] [42]
{kind=link}

A medida que los queratinocitos migran, se mueven sobre el tejido de granulación pero permanecen debajo de la costra, separando así la costra del tejido subyacente. [42] [47] Las células epiteliales tienen la capacidad de fagocitar desechos como tejido muerto y materia bacteriana que de otro modo obstruirían su camino. Debido a que deben disolver cualquier costra que se forme, la migración de queratinocitos se mejora mejor en un entorno húmedo, ya que uno seco conduce a la formación de una costra más grande y más resistente. [25] [37] [42] [50] Para abrirse camino a lo largo del tejido, los queratinocitos deben disolver el coágulo, los desechos y partes de la matriz extracelular para poder pasar. [47] [51] Secretan activador del plasminógeno , que activa el plasminógeno , convirtiéndolo en plasmina para disolver la costra. Las células sólo pueden migrar sobre tejido vivo, [42] por lo que deben excretar colagenasas y proteasas como las metaloproteinasas de matriz (MMP) para disolver las partes dañadas de la matriz extracelular en su camino, particularmente en la parte delantera de la capa migratoria. [47] Los queratinocitos también disuelven la membrana basal, utilizando en su lugar la nueva matriz extracelular depositada por los fibroblastos para atravesarla. [23]
A medida que los queratinocitos continúan migrando, se deben formar nuevas células epiteliales en los bordes de la herida para reemplazarlos y proporcionar más células para la capa que avanza. [25] La proliferación detrás de los queratinocitos migratorios normalmente comienza unos días después de la herida [46] y ocurre a una tasa que es 17 veces mayor en esta etapa de epitelización que en los tejidos normales. [25] Hasta que se renueva toda el área de la herida, las únicas células epiteliales que proliferan están en los bordes de la herida. [45]
Los factores de crecimiento, estimulados por las integrinas y las MMP, hacen que las células proliferen en los bordes de la herida. Los propios queratinocitos también producen y secretan factores, incluidos factores de crecimiento y proteínas de la membrana basal, que ayudan tanto en la epitelización como en otras fases de la curación. [52] Los factores de crecimiento también son importantes para la defensa inmunitaria innata de las heridas de la piel mediante la estimulación de la producción de péptidos antimicrobianos y citocinas quimiotácticas de neutrófilos en los queratinocitos.
Los queratinocitos continúan migrando a través del lecho de la herida hasta que las células de ambos lados se encuentran en el medio, momento en el que la inhibición por contacto hace que dejen de migrar. [31] Cuando han terminado de migrar, los queratinocitos secretan las proteínas que forman la nueva membrana basal. [31] Las células revierten los cambios morfológicos que experimentaron para comenzar a migrar; restablecen los desmosomas y hemidesmosomas y se anclan nuevamente a la membrana basal. [23] Las células basales comienzan a dividirse y diferenciarse de la misma manera que lo hacen en la piel normal para restablecer los estratos que se encuentran en la piel reepitelizada. [31]
Contracción
La contracción es una fase clave de la curación de heridas y su reparación. Si la contracción continúa durante demasiado tiempo, puede provocar desfiguración y pérdida de función. [32] Por lo tanto, existe un gran interés en comprender la biología de la contracción de las heridas, que se puede modelar in vitro utilizando el ensayo de contracción en gel de colágeno o el modelo dérmico equivalente . [27] [53]
La contracción comienza aproximadamente una semana después de la herida, cuando los fibroblastos se han diferenciado en miofibroblastos . [54] En heridas de espesor total, la contracción alcanza su punto máximo entre 5 y 15 días después de la herida. [37] La contracción puede durar varias semanas [46] y continúa incluso después de que la herida esté completamente reepitelizada. [3] Una herida grande puede volverse entre un 40 y un 80 % más pequeña después de la contracción. [31] [42] Las heridas pueden contraerse a una velocidad de hasta 0,75 mm por día, dependiendo de qué tan suelto esté el tejido en el área herida. [37] La contracción generalmente no ocurre simétricamente; más bien, la mayoría de las heridas tienen un "eje de contracción" que permite una mayor organización y alineación de las células con el colágeno. [54]
Al principio, la contracción se produce sin la participación de los miofibroblastos. [55] Más tarde, los fibroblastos, estimulados por factores de crecimiento, se diferencian en miofibroblastos. Los miofibroblastos, que son similares a las células musculares lisas, son responsables de la contracción. [55] Los miofibroblastos contienen el mismo tipo de actina que la que se encuentra en las células musculares lisas . [32]
Los miofibroblastos son atraídos por la fibronectina y los factores de crecimiento y se mueven a lo largo de la fibronectina unida a la fibrina en la matriz extracelular provisional para alcanzar los bordes de la herida. [25] Forman conexiones con la matriz extracelular en los bordes de la herida y se adhieren entre sí y a los bordes de la herida mediante desmosomas . Además, en una adhesión llamada fibronexo, la actina en el miofibroblasto se une a través de la membrana celular a moléculas en la matriz extracelular como la fibronectina y el colágeno. [55] Los miofibroblastos tienen muchas de estas adhesiones, que les permiten tirar de la matriz extracelular cuando se contraen, reduciendo el tamaño de la herida. [32] En esta parte de la contracción, el cierre se produce más rápidamente que en la primera parte, independiente de los miofibroblastos. [55]
A medida que la actina de los miofibroblastos se contrae, los bordes de la herida se juntan. Los fibroblastos depositan colágeno para reforzar la herida a medida que se contraen. [3] La etapa de contracción en la proliferación termina cuando los miofibroblastos dejan de contraerse y cometen apoptosis. [32] La degradación de la matriz provisional conduce a una disminución del ácido hialurónico y un aumento del sulfato de condroitina, lo que hace que los fibroblastos dejen de migrar y proliferar gradualmente. [19] Estos eventos señalan el inicio de la etapa de maduración de la cicatrización de la herida.
Maduración y remodelación
Cuando los niveles de producción y degradación de colágeno se igualan, se dice que ha comenzado la fase de maduración de la reparación tisular. [20] Durante la maduración, el colágeno tipo III , que prevalece durante la proliferación, es reemplazado por colágeno tipo I. [17] Las fibras de colágeno originalmente desorganizadas se reorganizan, se reticulan y se alinean a lo largo de las líneas de tensión . [31] El inicio de la fase de maduración puede variar ampliamente, dependiendo del tamaño de la herida y de si inicialmente se cerró o se dejó abierta, [28] desde aproximadamente tres días [41] hasta tres semanas. [56] La fase de maduración puede durar un año o más, de manera similar dependiendo del tipo de herida. [28]
A medida que avanza la fase, la resistencia a la tracción de la herida aumenta. [28] El colágeno alcanzará aproximadamente el 20% de su resistencia a la tracción después de tres semanas, aumentando al 80% después de 12 meses. La resistencia máxima de la cicatriz es el 80% de la de la piel sana. [57] Dado que la actividad en el sitio de la herida se reduce, la cicatriz pierde su apariencia roja a medida que los vasos sanguíneos que ya no son necesarios se eliminan por apoptosis . [20]
Las fases de cicatrización de heridas normalmente progresan de manera predecible y oportuna; si no es así, la curación puede progresar de manera inapropiada hasta convertirse en una herida crónica [7] como una úlcera venosa o en una cicatriz patológica como una cicatriz queloide . [58] [59]
Factores que afectan la cicatrización de heridas
Muchos factores que controlan la eficacia, la velocidad y la forma de cicatrización de las heridas se dividen en dos tipos: factores locales y sistémicos. [2]
Factores locales
- Humedad: mantener una herida húmeda en lugar de seca hace que la curación de la herida sea más rápida y con menos dolor y menos cicatrices [60]
- Factores mecánicos
- Edema
- Radiación ionizante
- Técnica defectuosa de cierre de heridas
- Isquemia y necrosis
- Cuerpos extraños. Los cuerpos extraños pequeños y afilados pueden penetrar la piel dejando una pequeña herida superficial pero causando lesiones internas y hemorragia interna. En el caso de un cuerpo extraño de vidrio, "con frecuencia, una herida inocente en la piel oculta la naturaleza extensa de las lesiones subyacentes". [61] La lesión nerviosa de primer grado requiere de unas horas a unas semanas para recuperarse. [62] Si un cuerpo extraño pasa por un nervio y causa una lesión nerviosa de primer grado durante la entrada, entonces la sensación del cuerpo extraño o el dolor debido a la herida interna puede demorarse unas horas o unas semanas después de la entrada. Un aumento repentino del dolor durante las primeras semanas de curación de la herida podría ser un signo de un nervio recuperado que informa lesiones internas en lugar de una infección recientemente desarrollada.
- Baja tensión de oxígeno
- Perfusión
Factores sistémicos
- Inflamación
- Diabetes : las personas con diabetes tienen una capacidad reducida para cicatrizar heridas agudas. Además, las personas diabéticas son propensas a desarrollar úlceras crónicas del pie diabético, una complicación grave de la diabetes que afecta al 15 % de las personas con diabetes y representa el 84 % de todas las amputaciones de la parte inferior de las piernas relacionadas con la diabetes. [63] La capacidad de cicatrización deteriorada de los diabéticos con úlceras del pie diabético o heridas agudas implica múltiples mecanismos fisiopatológicos. [64] Esta cicatrización deteriorada implica hipoxia, disfunción de fibroblastos y células epidérmicas, alteración de la angiogénesis y la neovascularización, altos niveles de metaloproteasas, daño por especies reactivas de oxígeno y AGE (productos finales de la glicación avanzada), disminución de la resistencia inmunitaria del huésped y neuropatía. [64]
- Nutrientes – La desnutrición o las deficiencias nutricionales tienen un impacto reconocible en la cicatrización de heridas después de un traumatismo o una intervención quirúrgica. [65] Los nutrientes, incluidas las proteínas, los carbohidratos, la arginina, la glutamina, los ácidos grasos poliinsaturados, la vitamina A, la vitamina C, la vitamina E, el magnesio, el cobre, el zinc y el hierro, desempeñan papeles importantes en la cicatrización de heridas. [64] Las grasas y los carbohidratos proporcionan la mayor parte de la energía necesaria para la cicatrización de heridas. La glucosa es la fuente de combustible más importante y se utiliza para crear ATP celular, proporcionando energía para la angiogénesis y la deposición de nuevos tejidos. [64] Como las necesidades nutricionales de cada paciente y su herida asociada son complejas, se sugiere que un apoyo nutricional personalizado beneficiaría tanto la cicatrización de heridas agudas como crónicas. [64]
- Enfermedades metabólicas
- Inmunosupresión
- Trastornos del tejido conectivo
- Fumar: el tabaquismo retrasa la curación de las heridas, sobre todo en las fases proliferativa e inflamatoria. También aumenta la probabilidad de que se produzcan ciertas complicaciones, como la rotura de la herida, la necrosis de la herida y del colgajo, la disminución de la resistencia a la tracción de la herida y la infección. [64] El tabaquismo pasivo también perjudica el proceso de cicatrización adecuado de las heridas. [66]
- Edad – La edad avanzada (más de 60 años) es un factor de riesgo para la cicatrización deficiente de las heridas. [64] Se reconoce que, en adultos mayores con buena salud en general, los efectos del envejecimiento causan un retraso temporal en la cicatrización, pero no un deterioro importante con respecto a la calidad de la cicatrización. [67] La cicatrización tardía de las heridas en pacientes de edad avanzada se asocia con una respuesta inflamatoria alterada; por ejemplo, una infiltración tardía de células T en la herida con alteraciones en la producción de quimiocinas y una capacidad fagocítica reducida de los macrófagos. [68]
- Alcohol: el consumo de alcohol perjudica la cicatrización de las heridas y también aumenta las posibilidades de infección. El alcohol afecta la fase proliferativa de la cicatrización. Una sola unidad de alcohol causa un efecto negativo en la reepitelización, el cierre de la herida, la producción de colágeno y la angiogénesis. [64]
En la década de 2000 surgieron los primeros modelos matemáticos del proceso de curación, basados en supuestos simplificados y en un sistema de ecuaciones diferenciales resueltas mediante MATLAB . Los modelos muestran que la "velocidad del proceso de curación" parece estar "muy influenciada por la actividad y el tamaño de la propia lesión, así como por la actividad del agente curativo". [69]
Investigación y desarrollo
Hasta aproximadamente el año 2000, el paradigma clásico de la cicatrización de heridas, que involucra células madre restringidas a linajes específicos de órganos, nunca había sido cuestionado seriamente. Desde entonces, la noción de que las células madre adultas tienen plasticidad celular o la capacidad de diferenciarse en células que no pertenecen a su linaje ha surgido como una explicación alternativa. [1] Para ser más específicos, las células progenitoras hematopoyéticas (que dan lugar a células maduras en la sangre) pueden tener la capacidad de desdiferenciarse nuevamente en células madre hematopoyéticas y/o transdiferenciarse en células que no pertenecen a su linaje, como los fibroblastos. [39]
Células madre y plasticidad celular
Las células madre adultas multipotentes tienen la capacidad de autorrenovarse y dar lugar a diferentes tipos de células. Las células madre dan lugar a células progenitoras, que son células que no se autorrenuevan, pero que pueden generar varios tipos de células. El grado de participación de las células madre en la cicatrización de heridas cutáneas (piel) es complejo y no se comprende del todo. [ cita requerida ] La inyección de células madre conduce a la cicatrización de heridas principalmente a través de la estimulación de la angiogénesis. [70]
Se cree que la epidermis y la dermis se reconstituyen mediante células madre mitóticamente activas que residen en el vértice de las crestas intercelulares (células madre basales o BSC), el bulto de los folículos pilosos (células madre foliculares pilosas o HFSC) y la dermis papilar (células madre dérmicas). [1] Además, la médula ósea también puede contener células madre que desempeñan un papel importante en la cicatrización de heridas cutáneas. [39]
En circunstancias raras, como una lesión cutánea extensa, las subpoblaciones de autorrenovación en la médula ósea se inducen a participar en el proceso de curación, por lo que dan lugar a células secretoras de colágeno que parecen desempeñar un papel durante la reparación de la herida. [1] Estas dos subpoblaciones de autorrenovación son (1) células madre mesenquimales derivadas de la médula ósea (MSC) y (2) células madre hematopoyéticas (HSC). La médula ósea también alberga una subpoblación progenitora ( células progenitoras endoteliales o EPC) que, en el mismo tipo de entorno, se movilizan para ayudar en la reconstrucción de los vasos sanguíneos. [39] Además, se cree que una lesión extensa en la piel también promueve el tráfico temprano de una subclase única de leucocitos ( fibrocitos circulantes ) a la región lesionada, donde realizan varias funciones relacionadas con la cicatrización de heridas. [1]
Reparación de heridas versus regeneración
Una lesión es una interrupción de la morfología y/o funcionalidad de un tejido determinado. Después de una lesión, el tejido estructural se cura con una regeneración incompleta o completa. [71] [72] El tejido sin una interrupción de la morfología casi siempre se regenera por completo. Un ejemplo de regeneración completa sin una interrupción de la morfología es el tejido no lesionado, como la piel. [73] La piel no lesionada tiene un reemplazo y regeneración continua de células que siempre da como resultado una regeneración completa. [73]
Existe una distinción sutil entre "reparación" y "regeneración". [1] [71] [72] Reparación significa regeneración incompleta . [71] Reparación o regeneración incompleta, se refiere a la adaptación fisiológica de un órgano después de una lesión en un esfuerzo por restablecer la continuidad sin tener en cuenta el reemplazo exacto del tejido perdido/dañado. [71] La verdadera regeneración tisular o regeneración completa , [72] se refiere al reemplazo del tejido perdido/dañado con una copia "exacta", de modo que tanto la morfología como la funcionalidad se restablezcan por completo. [72] Aunque después de una lesión los mamíferos pueden regenerarse completamente de forma espontánea, por lo general no lo hacen por completo. Un ejemplo de un tejido que se regenera por completo después de una interrupción de la morfología es el endometrio ; el endometrio después del proceso de descomposición a través del ciclo menstrual se cura con una regeneración completa. [73]
En algunos casos, después de una degradación tisular, como en la piel, se puede inducir una regeneración más cercana a la regeneración completa mediante el uso de andamios biodegradables ( colágeno - glicoaminoglicano ). Estos andamios son estructuralmente análogos a la matriz extracelular (ECM) que se encuentra en la dermis normal/no lesionada. [74] Las condiciones fundamentales requeridas para la regeneración tisular a menudo se oponen a las condiciones que favorecen la reparación eficiente de las heridas, incluida la inhibición de (1) la activación plaquetaria, (2) la respuesta inflamatoria y (3) la contracción de la herida. [1] Además de brindar apoyo para la adhesión de fibroblastos y células endoteliales, los andamios biodegradables inhiben la contracción de la herida, lo que permite que el proceso de curación avance hacia una vía más regenerativa/menos cicatricial. Se han investigado agentes farmacéuticos que pueden desactivar la diferenciación de miofibroblastos . [75]
Una nueva forma de pensar se deriva de la noción de que los heparán sulfatos son un elemento clave en la homeostasis tisular: el proceso que hace que el tejido reemplace las células muertas por células idénticas. En las zonas de heridas, la homeostasis tisular se pierde a medida que los heparán sulfatos se degradan, impidiendo el reemplazo de células muertas por células idénticas. Los análogos de heparán sulfato no pueden ser degradados por todas las heparanasas y glicanasas conocidas y se unen a los puntos de unión de heparín sulfato libre en la matriz extracelular, preservando así la homeostasis tisular normal y previniendo la cicatrización. [76] [77] [78]
Reparación o regeneración en relación con el factor inducible por hipoxia 1-alfa (HIF-1a). En circunstancias normales, después de una lesión, el HIF-1a es degradado por las prolil hidroxilasas (PHD). Los científicos descubrieron que la simple regulación positiva del HIF-1a a través de inhibidores de PHD regenera el tejido perdido o dañado en mamíferos que tienen una respuesta de reparación; y la regulación negativa continua del HIF-1a da como resultado la curación con una respuesta de cicatrización en mamíferos con una respuesta regenerativa previa a la pérdida de tejido. El acto de regular el HIF-1a puede activar o desactivar el proceso clave de la regeneración de los mamíferos. [79] [80]
Curación de heridas sin cicatrices
La curación de heridas sin cicatrices es un concepto basado en la curación o reparación de la piel (u otros tejidos u órganos) después de una lesión con el objetivo de curar con tejido cicatricial subjetivamente y relativamente menor del que se espera normalmente. La curación sin cicatrices a veces se confunde con el concepto de curación sin cicatrices , que es la curación de heridas que no deja cicatrices en absoluto . Sin embargo, son conceptos diferentes.
Un método inverso a la cicatrización de heridas sin cicatrices es la escarificación (la cicatrización de la herida deja más cicatrices). Históricamente, ciertas culturas consideran atractiva la escarificación; [81] sin embargo, este no suele ser el caso en la sociedad occidental moderna, en la que muchos pacientes acuden a las clínicas de cirugía plástica con expectativas poco realistas. Dependiendo del tipo de cicatriz , el tratamiento puede ser invasivo (inyecciones de esteroides intralesionales, cirugía) y/o conservador ( terapia de compresión , gel de silicona tópico , braquiterapia , terapia fotodinámica ). [82] Es necesario el juicio clínico para equilibrar con éxito los posibles beneficios de los diversos tratamientos disponibles frente a la probabilidad de una mala respuesta y las posibles complicaciones resultantes de estos tratamientos. Muchos de estos tratamientos pueden tener solo un efecto placebo , y la base de evidencia para el uso de muchos de los tratamientos actuales es pobre. [83]
Desde la década de 1960, la comprensión de los procesos biológicos básicos involucrados en la reparación de heridas y la regeneración de tejidos se ha expandido debido a los avances en biología celular y molecular . [84] Actualmente, los objetivos principales en el tratamiento de heridas son lograr un cierre rápido de la herida con un tejido funcional que tenga una cicatrización estética mínima. [85] Sin embargo, el objetivo final de la biología de la cicatrización de heridas es inducir una reconstrucción más perfecta del área de la herida. La cicatrización de heridas sin cicatrices solo ocurre en tejidos fetales de mamíferos [86] y la regeneración completa se limita a vertebrados inferiores, como salamandras e invertebrados . [87] En humanos adultos, el tejido lesionado se repara por deposición de colágeno , remodelación de colágeno y eventual formación de cicatrices, donde se cree que la cicatrización de heridas fetales es más un proceso regenerativo con mínima o ninguna formación de cicatrices. [86] Por lo tanto, la cicatrización de heridas fetales se puede utilizar para proporcionar un modelo mamífero accesible de una respuesta de curación óptima en tejidos humanos adultos. Las pistas sobre cómo podría lograrse esto provienen de estudios de cicatrización de heridas en embriones, donde la reparación es rápida y eficiente y da como resultado una regeneración esencialmente perfecta de cualquier tejido perdido.
La etimología del término curación de heridas sin cicatrices tiene una larga historia. [88] [89] [90] El concepto anticuado de curación sin cicatrices se presentó en forma impresa a principios del siglo XX y apareció en un artículo publicado en la revista London Lancet. Este proceso implicaba cortar en una inclinación quirúrgica respecto a la superficie de la piel, en lugar de en un ángulo recto; el proceso se describió en varios periódicos. [88] [89] [90]
Cáncer
Después de la inflamación, la restauración de la integridad y la función normal del tejido se preserva mediante interacciones de retroalimentación entre diversos tipos de células mediadas por moléculas de adhesión y citocinas secretadas. La alteración de los mecanismos de retroalimentación normales en el cáncer amenaza la integridad del tejido y permite que un tumor maligno escape al sistema inmunológico. [91] [92] Un ejemplo de la importancia de la respuesta de curación de heridas en los tumores se ilustra en el trabajo de Howard Chang y sus colegas de la Universidad de Stanford que estudian los cánceres de mama. [8]
Suplementos orales de colágeno
Los resultados preliminares son prometedores para el uso a corto y largo plazo de suplementos orales de colágeno para la cicatrización de heridas y el envejecimiento de la piel. Los suplementos orales de colágeno también aumentan la elasticidad de la piel, la hidratación y la densidad del colágeno dérmico. La suplementación con colágeno es generalmente segura y no se han reportado efectos adversos. Se necesitan más estudios para dilucidar el uso médico en enfermedades de la barrera cutánea como la dermatitis atópica y para determinar los regímenes de dosificación óptimos. [93]
Vendajes para heridas
En los últimos años se han llevado a cabo numerosas investigaciones y se han desarrollado apósitos modernos para ayudar a la curación de heridas. Los científicos se proponen desarrollar apósitos para heridas que tengan las siguientes características: [94]
- Proporcionar protección contra heridas
- Eliminar el exceso de exudado
- Poseen propiedades antimicrobianas
- Mantener un ambiente húmedo
- Tienen alta permeabilidad al oxígeno.
- Se eliminan fácilmente de la zona de la herida.
- Poseen características no anafilácticas.
Los apósitos de gasa de algodón han sido el estándar de atención, a pesar de sus propiedades secas que pueden adherirse a las superficies de las heridas y causar molestias al retirarlos. Investigaciones recientes se han propuesto mejorar los apósitos de gasa de algodón para acercarlos a las propiedades de los apósitos para heridas modernos, recubriendo el apósito de gasa de algodón para heridas con un nanocompuesto de quitosano / Ag / ZnO . Estos apósitos actualizados proporcionan una mayor absorción de agua y una eficacia antibacteriana mejorada . [94]
Limpieza de heridas
Se puede limpiar la suciedad o el polvo de la superficie de la herida, las bacterias, el tejido que ha muerto y el líquido de la herida. La evidencia que respalda la técnica más eficaz no es clara y no hay evidencia suficiente para concluir si la limpieza de las heridas es beneficiosa para promover la curación o si las soluciones de limpieza de heridas ( polihexametilen biguanida , peróxido de hidrógeno acuoso , etc.) son mejores que el agua esterilizada o las soluciones salinas para ayudar a curar las úlceras venosas de las piernas. [95] No se sabe con certeza si la elección de la solución de limpieza o el método de aplicación influyen en la curación de las úlceras venosas de las piernas . [95]
Simulación de la cicatrización de heridas desde una perspectiva de crecimiento
Se ha dedicado un esfuerzo considerable a comprender las relaciones físicas que rigen la curación de las heridas y la posterior formación de cicatrices, y se han desarrollado modelos matemáticos y simulaciones para dilucidar estas relaciones. [96] El crecimiento de tejido alrededor del sitio de la herida es el resultado de la migración de células y la deposición de colágeno por parte de estas células. La alineación del colágeno describe el grado de cicatrización; la orientación en forma de tejido de canasta del colágeno es característica de la piel normal, mientras que las fibras de colágeno alineadas dan lugar a una cicatrización significativa. [97] Se ha demostrado que el crecimiento de tejido y la extensión de la formación de cicatrices se pueden controlar modulando la tensión en el sitio de la herida. [98]
El crecimiento de los tejidos se puede simular utilizando las relaciones antes mencionadas desde un punto de vista bioquímico y biomecánico. Los productos químicos biológicamente activos que desempeñan un papel importante en la cicatrización de heridas se modelan con difusión fickiana para generar perfiles de concentración. La ecuación de equilibrio para sistemas abiertos al modelar la cicatrización de heridas incorpora el crecimiento de la masa debido a la migración y proliferación celular. Aquí se utiliza la siguiente ecuación:
D t ρ 0 = Div (R) + R 0 ,
donde ρ representa la densidad de masa, R representa un flujo de masa (de la migración celular) y R 0 representa una fuente de masa (de la proliferación, división o agrandamiento celular). [99] Relaciones como estas se pueden incorporar en modelos basados en agentes , donde se puede probar la sensibilidad a parámetros individuales como la alineación inicial del colágeno, las propiedades de las citocinas y las tasas de proliferación celular. [100]
Intenciones de cierre de heridas
La cicatrización exitosa de heridas depende de varios tipos de células, mediadores moleculares y elementos estructurales. [101]
Intención primaria
La intención principal es la curación de una herida limpia sin pérdida de tejido. [101] En este proceso, los bordes de la herida se juntan, de modo que queden adyacentes entre sí (reaproximados). El cierre de la herida se realiza con suturas (puntos), grapas o cinta adhesiva o pegamento.
La intención primaria sólo se puede implementar cuando la herida es precisa y hay una alteración mínima del tejido local y de la membrana basal epitelial, por ejemplo, incisiones quirúrgicas. [102]
Este proceso es más rápido que la curación por segunda intención. [101] También hay menos cicatrices asociadas con la primera intención, ya que no hay grandes pérdidas de tejido que rellenar con tejido de granulación, aunque se formará algo de tejido de granulación. [101]
- Los ejemplos de intención primaria incluyen: laceraciones bien reparadas , fracturas óseas bien reducidas y curación después de una cirugía de colgajo .
- La retirada temprana de los apósitos de heridas limpias o limpias-contaminadas afecta la cicatrización primaria de las heridas. [103] [ se necesita una mejor fuente ]
Intención secundaria
- La intención secundaria se implementa cuando la intención primaria no es posible debido a un daño o pérdida significativa de tejido, generalmente debido a que la herida fue creada por un traumatismo importante. [102]
- Se deja que la herida se granule.
- El cirujano puede tapar la herida con una gasa o utilizar un sistema de drenaje.
- La granulación produce una cicatriz más ancha.
- El proceso de curación puede ser lento debido a la presencia de drenaje de la infección.
- El cuidado de la herida debe realizarse diariamente para estimular la eliminación de restos de la herida y permitir la formación de tejido de granulación.
- El uso de antibióticos o antisépticos para la cicatrización de heridas quirúrgicas por segunda intención es controvertido. [104]
- Ejemplos: gingivectomía , gingivoplastia , alvéolos para extracción dentaria , fracturas mal reducidas, quemaduras, laceraciones graves, úlceras por presión.
- No hay evidencia suficiente de que la elección de apósitos o agentes tópicos afecte la cicatrización secundaria de las heridas. [105]
- No existe evidencia de la efectividad de la terapia de presión negativa en la cicatrización de heridas por segunda intención. [106]
Intención terciaria
(Cierre primario retardado):
- La herida se limpia, se desbrida y se observa inicialmente, normalmente 4 o 5 días antes del cierre.
- La herida se deja abierta intencionadamente.
- Ejemplos: curación de heridas mediante el uso de injertos de tejido .
Si los bordes de la herida no se vuelven a aproximar inmediatamente, se produce un retraso en la cicatrización primaria de la herida. Este tipo de cicatrización puede ser deseable en el caso de heridas contaminadas. Al cuarto día, la fagocitosis de los tejidos contaminados ya está en marcha y se están produciendo los procesos de epitelización, deposición de colágeno y maduración. Los materiales extraños son bloqueados por macrófagos que pueden metamorfosearse en células epitelioides, que están rodeadas por leucocitos mononucleares, formando granulomas. Por lo general, la herida se cierra quirúrgicamente en este momento, o se come la costra, y si la "limpieza" de la herida es incompleta, puede producirse una inflamación crónica, lo que da lugar a una cicatrización prominente.
Descripción general de los factores de crecimiento involucrados
Los siguientes son los principales factores de crecimiento que intervienen en la cicatrización de heridas:
| Factor de crecimiento | Abreviatura | Orígenes principales | Efectos |
|---|---|---|---|
| Factor de crecimiento epidérmico | FCE |
| |
| Factor de crecimiento transformante-α | TGF-α |
|
|
| Factor de crecimiento de hepatocitos | factor de crecimiento humano |
| |
| Factor de crecimiento endotelial vascular | Factor de crecimiento endotelial vascular (VEGF) |
|
|
| Factor de crecimiento derivado de plaquetas | PDGF |
|
|
| Factor de crecimiento de fibroblastos 1 y 2 | FGF-1, −2 |
|
|
| Factor de crecimiento transformante β | TGF-β |
|
|
| Factor de crecimiento de queratinocitos | KGF |
|
|
| A menos que se especifique lo contrario en los recuadros, la referencia es: [107] | |||
Complicaciones de la cicatrización de heridas
Las principales complicaciones son muchas:
- Formación deficiente de cicatrices: produce dehiscencia de la herida o ruptura de la herida debido a una formación inadecuada de tejido de granulación.
- Formación excesiva de cicatrices: cicatriz hipertrófica , queloide , desmoide .
- Granulación exuberante ( carne orgullosa ).
- Contracción deficiente (en injertos de piel) o contracción excesiva (en quemaduras).
- Otros: Calcificación distrófica , cambios pigmentarios, cicatrices dolorosas, hernia incisional.
Otras complicaciones pueden incluir infección y úlcera de Marjolin .
Productos biológicos, sustitutos de la piel, biomembranas y andamios
Los avances en la comprensión clínica de las heridas y su fisiopatología han dado lugar a importantes innovaciones biomédicas en el tratamiento de heridas agudas, crónicas y de otros tipos. Se han desarrollado numerosos productos biológicos, sustitutos de la piel , biomembranas y andamios para facilitar la cicatrización de heridas a través de diversos mecanismos. [108] Esto incluye una serie de productos con nombres comerciales como Epicel , Laserskin , Transcyte, Dermagraft, AlloDerm/Strattice, Biobrane, Integra, Apligraf, OrCel, GraftJacket y PermaDerm. [109]
Véase también
Notas y referencias
- ^ abcdefghijkl Nguyen DT, Orgill DP, Murphy GT (2009). "4 La base fisiopatológica de la cicatrización de heridas y la regeneración cutánea". En Orgill DP, Blanco C (eds.). Biomateriales para el tratamiento de la pérdida de piel . Elsevier. págs. 25–57. ISBN 978-1-84569-554-5.
- ^ ab Rieger S, Zhao H, Martin P, Abe K, Lisse TS (enero de 2015). "El papel de los receptores nucleares de hormonas en la reparación de heridas cutáneas". Cell Biochemistry and Function . 33 (1): 1–13. doi :10.1002/cbf.3086. PMC 4357276 . PMID 25529612.
- ^ abcdefghijkl Stadelmann WK, Digenis AG, Tobin GR (agosto de 1998). "Fisiología y dinámica de curación de heridas cutáneas crónicas". American Journal of Surgery . 176 (2A Suppl): 26S–38S. doi :10.1016/S0002-9610(98)00183-4. PMID 9777970.
- ^ Enoch, S. Price, P. (2004). Diferencias celulares, moleculares y bioquímicas en la fisiopatología de la curación entre heridas agudas, heridas crónicas y heridas en ancianos Archivado el 6 de julio de 2017 en Wayback Machine .
- ^ Rasche H (2001). "Hemóstasis y trombosis: una descripción general". Suplementos del European Heart Journal . 3 (Suplemento Q): Q3–Q7. doi : 10.1016/S1520-765X(01)90034-3 .
- ^ Versteeg HH, Heemskerk JW, Levi M, Reitsma PH (enero de 2013). "Nuevos fundamentos en hemostasia". Physiological Reviews . 93 (1): 327–58. doi :10.1152/physrev.00016.2011. PMID 23303912. S2CID 10367343.
- ^ abcdefgh Midwood KS, Williams LV, Schwarzbauer JE (junio de 2004). "Reparación tisular y dinámica de la matriz extracelular". Revista internacional de bioquímica y biología celular . 36 (6): 1031–7. doi :10.1016/j.biocel.2003.12.003. PMID 15094118.
- ^ ab Chang HY, Sneddon JB, Alizadeh AA, Sood R, West RB, Montgomery K, et al. (febrero de 2004). "La firma de expresión génica de la respuesta sérica de fibroblastos predice la progresión del cáncer humano: similitudes entre tumores y heridas". PLOS Biology . 2 (2): E7. doi : 10.1371/journal.pbio.0020007 . PMC 314300 . PMID 14737219.
- ^ Garg, HG (2000). Cicatrización de heridas sin cicatrices . Nueva York Marcel Dekker, Inc. Libro electrónico.
- ^ La lista de referencias se encuentra en la página principal de la imagen .
- ^ Cubison TC, Pape SA, Parkhouse N (diciembre de 2006). "Evidencia de la relación entre el tiempo de curación y el desarrollo de cicatrices hipertróficas (HTS) en quemaduras pediátricas debidas a lesiones por escaldadura". Burns . 32 (8): 992–9. doi :10.1016/j.burns.2006.02.007. PMID 16901651.
- ^ Kraft J, Lynde C. "Quemaduras de primer, segundo y tercer grado: clasificación de las quemaduras". skincareguide.ca. Archivado desde el original el 26 de diciembre de 2011. Consultado el 31 de enero de 2012.
Formación de una escara gruesa, curación lenta (>1 mes), cicatrización evidente,
- ^ ab "CICATRIZ POST QUEMADURA EN RELACIÓN CON LA REEPITELIZACIÓN". Burnsurgery.org. 2011. Archivado desde el original el 25 de abril de 2012. Consultado el 16 de marzo de 2011. Curación en 2 semanas: cicatriz mínima o nula; Curación en 3 semanas: cicatriz mínima o nula ,
excepto en pacientes con alto riesgo de formación de cicatrices; Curación en 4 semanas o más: hipertrófica en más del 50 % de los pacientes
- ^ Galko MJ, Krasnow MA (agosto de 2004). "Análisis celular y genético de la cicatrización de heridas en larvas de Drosophila". PLOS Biology . 2 (8): E239. doi : 10.1371/journal.pbio.0020239 . PMC 479041 . PMID 15269788.
- ^ abcde Rosenberg L., de la Torre J. (2006). Cicatrización de heridas, factores de crecimiento Archivado el 21 de noviembre de 2008 en Wayback Machine . Emedicine.com. Consultado el 20 de enero de 2008.
- ^ Sandeman SR, Allen MC, Liu C, Faragher RG, Lloyd AW (noviembre de 2000). "La migración de queratocitos humanos a geles de colágeno disminuye con el envejecimiento in vitro". Mecanismos de envejecimiento y desarrollo . 119 (3): 149–57. doi :10.1016/S0047-6374(00)00177-9. PMID 11080534. S2CID 21115977.
- ^ abcd Dealey C. (1999). El cuidado de las heridas: una guía para enfermeras . Oxford; Malden, Mass. Blackwell Science. Libro electrónico.
- ^ Theoret CL (2004). "Actualización en la reparación de heridas". Técnicas Clínicas en la Práctica Equina . 3 (2): 110–122. doi :10.1053/j.ctep.2004.08.009.
- ^ abcdefgh de la Torre J., Sholar A. (2006). Cicatrización de heridas: heridas crónicas Archivado el 29 de octubre de 2008 en Wayback Machine . Emedicine.com. Consultado el 20 de enero de 2008.
- ^ abcdefgh Greenhalgh DG (septiembre de 1998). "El papel de la apoptosis en la cicatrización de heridas". Revista internacional de bioquímica y biología celular . 30 (9): 1019–30. doi :10.1016/S1357-2725(98)00058-2. PMID 9785465.
- ^ Muller MJ, Hollyoak MA, Moaveni Z, Brown TL, Herndon DN, Heggers JP (diciembre de 2003). "El retraso de la cicatrización de heridas causado por la sulfadiazina de plata se revierte con aloe vera y nistatina". Burns . 29 (8): 834–6. doi :10.1016/S0305-4179(03)00198-0. PMID 14636760.
- ^ Martin P, Leibovich SJ (noviembre de 2005). "Células inflamatorias durante la reparación de heridas: lo bueno, lo malo y lo feo". Tendencias en biología celular . 15 (11): 599–607. doi :10.1016/j.tcb.2005.09.002. PMID 16202600.
- ^ abcdefgh Santoro MM, Gaudino G (marzo de 2005). "Facetas celulares y moleculares de la reepitelización de los queratinocitos durante la cicatrización de heridas". Experimental Cell Research . 304 (1): 274–86. doi :10.1016/j.yexcr.2004.10.033. PMID 15707592.
- ^ ab "Las fases de la cicatrización de heridas cutáneas" (PDF) . Expert Reviews in Molecular Medicine . 5 . Cambridge University Press. 21 de marzo de 2003. Archivado desde el original (PDF) el 8 de marzo de 2008.
- ^ abcdef Deodhar AK, Rana RE (1997). "Fisiología quirúrgica de la cicatrización de heridas: una revisión". Revista de Medicina de Postgrado . 43 (2): 52–6. PMID 10740722. Archivado desde el original el 26 de febrero de 2011. Consultado el 27 de octubre de 2005 .
- ^ Ovchinnikov DA (septiembre de 2008). "Macrófagos en el embrión y más allá: mucho más que fagocitos gigantes". Génesis . 46 (9): 447–62. doi : 10.1002/dvg.20417 . PMID 18781633. S2CID 38894501.
Los macrófagos están presentes esencialmente en todos los tejidos, comenzando con el desarrollo embrionario y, además de su papel en la defensa del huésped y en la eliminación de células apoptóticas, cada vez se reconocen más por su función trófica y su papel en la regeneración.
- ^ abcd Newton PM, Watson JA, Wolowacz RG, Wood EJ (agosto de 2004). "Los macrófagos restringen la contracción de un modelo de cicatrización de heridas in vitro". Inflamación . 28 (4): 207–14. doi :10.1023/B:IFLA.0000049045.41784.59. PMID 15673162. S2CID 9612298.
- ^ abcdefgh Mercandetti M, Cohen AJ (2005). "Cicatrización de heridas: curación y reparación". Emedicine.com . Archivado desde el original el 21 de noviembre de 2008. Consultado el 20 de enero de 2008 .
- ^ Swirski FK, Nahrendorf M, Etzrodt M, Wildgruber M, Cortez-Retamozo V, Panizzi P, Figueiredo JL, Kohler RH, et al. (julio de 2009). "Identificación de monocitos de reservorio esplénico y su despliegue en sitios inflamatorios". Science . 325 (5940): 612–6. Bibcode :2009Sci...325..612S. doi :10.1126/science.1175202. PMC 2803111 . PMID 19644120.
- ^ Jia T, Pamer EG (julio de 2009). "Inmunología. Prescindible pero no irrelevante". Science . 325 (5940): 549–50. Bibcode :2009Sci...325..549J. doi :10.1126/science.1178329. PMC 2917045 . PMID 19644100.
- ^ abcdefghi Lorenz HP, Longaker MT (2008). "Heridas: biología, patología y tratamiento" (PDF) . En Norton JA (ed.). Cirugía . Nueva York, NY: Springer. págs. 191–208. doi :10.1007/978-0-387-68113-9_10. ISBN 978-0-387-30800-5. S2CID 83849346. Archivado desde el original (PDF) el 24 de agosto de 2014.
- ^ abcde Hinz B (abril de 2006). "Maestros y sirvientes de la fuerza: el papel de las adhesiones a la matriz en la percepción y transmisión de la fuerza de los miofibroblastos". Revista Europea de Biología Celular . 85 (3–4): 175–81. doi :10.1016/j.ejcb.2005.09.004. PMID 16546559.
- ^ Souppouris A (23 de mayo de 2013). "Los científicos identifican una célula que podría contener el secreto de la regeneración de las extremidades". the verge.com. Archivado desde el original el 31 de julio de 2017. Consultado el 18 de septiembre de 2017. Los
investigadores han identificado una célula que ayuda a la regeneración de las extremidades en las salamandras. Los macrófagos son un tipo de célula reparadora que devora células muertas y patógenos, y activa otras células inmunes para responder a los patógenos.
- ^ Godwin JW, Pinto AR, Rosenthal NA (junio de 2013). "Los macrófagos son necesarios para la regeneración de las extremidades de las salamandras adultas". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 110 (23). Universidad de Texas: 9415–20. Bibcode :2013PNAS..110.9415G. doi : 10.1073/pnas.1300290110 . PMC 3677454 . PMID 23690624.
- ^ abcd Falanga V. (2005). Cicatrización de heridas. Academia Estadounidense de Dermatología (AAD).
- ^ abc Kuwahara RT y Rasberry R. 2007. Peeling químico. Archivado el 25 de octubre de 2008 en Wayback Machine . Emedicine.com. Consultado el 15 de septiembre de 2007.
- ^ abcdefghij Romo T. y Pearson JM 2005. Cicatrización de heridas, piel Archivado el 7 de diciembre de 2008 en Wayback Machine . Emedicine.com. Consultado el 27 de diciembre de 2006.
- ^ Lansdown AB, Sampson B, Rowe A (febrero de 2001). "Observaciones experimentales en ratas sobre la influencia del cadmio en la reparación de heridas cutáneas". Revista internacional de patología experimental . 82 (1): 35–41. doi :10.1046/j.1365-2613.2001.00180.x. PMC 2517695 . PMID 11422539.
- ^ abcd Song G, Nguyen DT, Pietramaggiori G, Scherer S, Chen B, Zhan Q, Ogawa R, Yannas IV , Wagers AJ, Orgill DP, Murphy GF (2010). "Uso del modelo parabiótico en estudios de cicatrización de heridas cutáneas para definir la participación de las células circulantes". Reparación y regeneración de heridas . 18 (4): 426–32. doi :10.1111/j.1524-475X.2010.00595.x. PMC 2935287 . PMID 20546556.
- ^ Ruszczak Z (noviembre de 2003). "Efecto de las matrices de colágeno en la cicatrización de heridas dérmicas". Advanced Drug Delivery Reviews . 55 (12): 1595–611. doi :10.1016/j.addr.2003.08.003. PMID 14623403.
- ^ ab Fig. 9-1. Fases celular, bioquímica y mecánica de la cicatrización de heridas. Pollock RE, Brunicardi FC, Andersen DK, Billiar TR, Dunn D, Hunter JG, Matthews JJ (2009). Schwartz's Principles of Surgery, novena edición . McGraw-Hill Professional. ISBN 978-0-07-154769-7.
- ^ abcdefgh DiPietro LA, Burns AL, eds. (2003). Cicatrización de heridas: métodos y protocolos . Métodos en medicina molecular. Totowa, NJ: Humana Press.
- ^ Fu XB, Sun TZ, Li XK, Sheng ZY (febrero de 2005). "Características morfológicas y de distribución de las glándulas sudoríparas en cicatrices hipertróficas y sus posibles efectos en la regeneración de las glándulas sudoríparas". Revista Médica China . 118 (3): 186–91. PMID 15740645. Archivado desde el original el 2018-06-20 . Consultado el 18 de junio de 2014 .
- ^ ab "LESIONES POR QUEMADURAS". nationaltraumainstitute.org. Archivado desde el original el 3 de marzo de 2016. Consultado el 13 de julio de 2016. Cuando
se destruye la dermis, las cicatrices no permiten que vuelvan a crecer cabello, nervios ni glándulas sudoríparas, lo que plantea desafíos adicionales para el control de la temperatura corporal.
- ^ abcd Bartkova J, Grøn B, Dabelsteen E, Bartek J (febrero de 2003). "Proteínas reguladoras del ciclo celular en la cicatrización de heridas humanas". Archivos de biología oral . 48 (2): 125–32. doi :10.1016/S0003-9969(02)00202-9. PMID 12642231.
- ^ abcd Mulvaney M. y Harrington A. 1994. Capítulo 7: Traumatismo cutáneo y su tratamiento. En, Textbook of Military Medicine: Military Dermatology . Oficina del Cirujano General, Departamento del Ejército. Proyecto de Hospital Naval Virtual. Consultado a través del archivo web el 15 de septiembre de 2007.
- ^ abcd Larjava H., Koivisto L. y Hakkinen L. 2002. Capítulo 3: Interacciones de los queratinocitos con la fibronectina durante la cicatrización de heridas. En, Heino, J. y Kahari, VM Cell Invasion . Medical Intelligence Unit; 33. Georgetown, Tex., Austin, Tex Landes Bioscience, Inc. Libro electrónico.
- ^ Witte MB, Barbul A (abril de 2002). "El papel del óxido nítrico en la reparación de heridas". American Journal of Surgery . 183 (4): 406–12. doi :10.1016/S0002-9610(02)00815-2. PMID 11975928.
- ^ Son HJ, Bae HC, Kim HJ, Lee DH, Han DW, Park JC (2005). "Efectos del β-glucano en la proliferación y migración de fibroblastos". Current Applied Physics . 5 (5): 468–71. Bibcode :2005CAP.....5..468S. doi :10.1016/j.cap.2005.01.011.
- ^ Falanga V (2004). "La herida crónica: problemas de cicatrización y soluciones en el contexto de la preparación del lecho de la herida". Células sanguíneas, moléculas y enfermedades . 32 (1): 88–94. doi :10.1016/j.bcmd.2003.09.020. PMID 14757419.
- ^ Etscheid M, Beer N, Dodt J (diciembre de 2005). "La proteasa que se une al hialuronano regula positivamente las vías de señalización de ERK1/2 y PI3K/Akt en fibroblastos y estimula la proliferación y migración celular". Cellular Signalling . 17 (12): 1486–94. doi :10.1016/j.cellsig.2005.03.007. PMID 16153533.
- ^ Bayram Y, Deveci M, Imirzalioglu N, Soysal Y, Sengezer M (octubre de 2005). "El apósito basado en células con queratinocitos alogénicos vivos en el tratamiento de las úlceras del pie: un estudio de caso". British Journal of Plastic Surgery . 58 (7): 988–96. doi : 10.1016/j.bjps.2005.04.031 . PMID 16040019.
- ^ Grinnell F (febrero de 1994). "Fibroblastos, miofibroblastos y contracción de heridas". The Journal of Cell Biology . 124 (4): 401–4. doi :10.1083/jcb.124.4.401. PMC 2119916 . PMID 8106541.
- ^ ab Eichler MJ, Carlson MA (febrero de 2006). "Modelado del tejido de granulación dérmica con la matriz de colágeno lineal poblada por fibroblastos: una comparación con el modelo de matriz redonda". Journal of Dermatological Science . 41 (2): 97–108. doi :10.1016/j.jdermsci.2005.09.002. PMID 16226016.
- ^ abcd Mirastschijski U, Haaksma CJ, Tomasek JJ, Agren MS (octubre de 2004). "El inhibidor de metaloproteinasa de matriz GM 6001 atenúa la migración y contracción de queratinocitos y la formación de miofibroblastos en heridas de la piel". Experimental Cell Research . 299 (2): 465–75. doi :10.1016/j.yexcr.2004.06.007. PMID 15350544.
- ^ worldwidewounds.com Archivado el 5 de julio de 2011 en Wayback Machine > Figura 3: Relación temporal entre los diferentes procesos de cicatrización de heridas. Archivado el 18 de julio de 2011 en Wayback Machine por Gregory S Schultz, Glenn Ladwig y Annette Wysocki, a su vez adaptado de Asmussen PD, Sollner B. Mecanismo de cicatrización de heridas. En: Wound Care. Tutorial Medical Series. Stuttgart: Hippokrates Verlag, 1993.
- ^ Morton LM, Phillips TJ (abril de 2016). "Cicatría y tratamiento de heridas: diagnóstico diferencial y evaluación de heridas crónicas". Revista de la Academia Estadounidense de Dermatología . 74 (4): 589–605, cuestionario 605–6. doi :10.1016/j.jaad.2015.08.068. PMID 26979352.
- ^ O'Leary R, Wood EJ, Guillou PJ (2002). "Cicatriz patológica: intervenciones estratégicas". Revista Europea de Cirugía = Acta Chirurgica . 168 (10): 523–34. PMID 12666691.
- ^ Desmoulière A, Chaponnier C, Gabbiani G (2005). "Reparación tisular, contracción y miofibroblasto". Reparación y regeneración de heridas . 13 (1): 7–12. doi :10.1111/j.1067-1927.2005.130102.x. PMID 15659031. S2CID 2590702.
- ^ Metzger S (septiembre de 2004). "Ventajas clínicas y financieras del tratamiento húmedo de heridas". Enfermera de atención médica domiciliaria . 22 (9): 586–90. doi :10.1097/00004045-200409000-00003. PMID 15359168.
- ^ Iconomou TG, Zuker RM, Michelow BJ (1993). "Manejo de lesiones penetrantes de vidrio importantes en las extremidades superiores de niños y adolescentes". Microcirugía . 14 (2): 91–6. doi :10.1002/micr.1920140202. PMID 8469109. S2CID 25492817.
- ^ "Lesión nerviosa". Johns Hopkins Medicine . The Johns Hopkins University, The Johns Hopkins Hospital y Johns Hopkins Health System. Archivado desde el original el 27 de septiembre de 2016. Consultado el 2 de octubre de 2016 .
- ^ Brem H, Tomic-Canic M (mayo de 2007). "Bases celulares y moleculares de la cicatrización de heridas en la diabetes". The Journal of Clinical Investigation . 117 (5): 1219–22. doi :10.1172/jci32169. PMC 1857239 . PMID 17476353.
- ^ abcdefgh Guo S, Dipietro LA (marzo de 2010). "Factores que afectan la cicatrización de heridas". Revista de investigación dental . 89 (3): 219–29. doi :10.1177/0022034509359125. PMC 2903966 . PMID 20139336.
- ^ Arnold M, Barbul A (junio de 2006). "Nutrición y cicatrización de heridas". Cirugía Plástica y Reconstructiva . 117 (7 Suppl): 42S–58S. doi :10.1097/01.prs.0000225432.17501.6c. PMID 16799374. S2CID 8658373.
- ^ Wong LS, Green HM, Feugate JE, Yadav M, Nothnagel EA, Martins-Green M (abril de 2004). "Efectos del humo de segunda mano en la estructura y función de los fibroblastos, células que son fundamentales para la reparación y remodelación de tejidos". BMC Cell Biology . 5 (1): 13. doi : 10.1186/1471-2121-5-13 . PMC 400727 . PMID 15066202.
- ^ Gosain A, DiPietro LA (marzo de 2004). "Envejecimiento y cicatrización de heridas". Revista Mundial de Cirugía . 28 (3): 321–6. doi :10.1007/s00268-003-7397-6. PMID 14961191. S2CID 28491255.
- ^ Swift ME, Burns AL, Gray KL, DiPietro LA (noviembre de 2001). "Alteraciones relacionadas con la edad en la respuesta inflamatoria a la lesión dérmica". The Journal of Investigative Dermatology . 117 (5): 1027–35. doi : 10.1046/j.0022-202x.2001.01539.x . PMID 11710909.
- ^ Alfredo Palomino I.; Leighton Estrada R.; Javier Valeriano M.; Sergio Luque M. (24 de septiembre de 2019). "Modelamiento matemático del proceso de curación de heridas bajo la acción de un ingrediente farmacéuticamente activo (IFA). Selecciones Matemáticas (en inglés americano y español). 6 (2): 283–288. doi : 10.17268/sel.mat.2019.02.14 . ISSN 2411-1783. OCLC 8469127433. Archivado desde el original el 22 de julio de 2020 . Consultado el 29 de agosto de 2020 .
- ^ Krasilnikova, OA; Baranovskii, DS; Lyundup, AV; Shegay, PV; Kaprin, AD; Klabukov, ID (27 de abril de 2022). "Monoterapia con células madre y somáticas para el tratamiento de las úlceras del pie diabético: revisión de estudios clínicos y mecanismos de acción". Reseñas e informes de células madre . 18 (6): 1974–1985. doi :10.1007/s12015-022-10379-z. ISSN 2629-3277. PMID 35476187. S2CID 248402820.
- ^ abcd Min S, Wang SW, Orr W (2006). "Patología general gráfica: 2.3 Regeneración incompleta". Patología . pathol.med.stu.edu.cn. Archivado desde el original el 2013-11-10 . Consultado el 2012-12-07 .
El tejido nuevo no es el mismo que el tejido que se perdió. Una vez completado el proceso de reparación, se produce una pérdida en la estructura o función del tejido lesionado. En este tipo de reparación, es común que el tejido de granulación (tejido conectivo del estroma) prolifere para rellenar el defecto creado por las células necróticas. Las células necróticas son luego reemplazadas por tejido cicatricial.
- ^ abcd Min S, Wang SW, Orr W (2006). "Patología general gráfica: 2.2 regeneración completa". Patología . pathol.med.stu.edu.cn. Archivado desde el original el 2012-12-07 . Consultado el 2012-12-07 .
(1) Regeneración completa: el tejido nuevo es el mismo que el tejido que se perdió. Una vez completado el proceso de reparación, la estructura y la función del tejido dañado son completamente normales.
- ^ abc Min S, Wang SW, Orr W (2006). "Patología general gráfica: 2.2 regeneración completa". Patología . pathol.med.stu.edu.cn. Archivado desde el original el 2012-12-07 . Consultado el 2013-11-10 .
Una vez completado el proceso de reparación, la estructura y la función del tejido lesionado son completamente normales. Este tipo de regeneración es común en situaciones fisiológicas. Ejemplos de regeneración fisiológica son el reemplazo continuo de células de la piel y la reparación del endometrio después de la menstruación. La regeneración completa puede ocurrir en situaciones patológicas en tejidos que tienen una buena capacidad regenerativa.
- ^ Yannas IV, Lee E, Orgill DP, Skrabut EM, Murphy GF (febrero de 1989). "Síntesis y caracterización de una matriz extracelular modelo que induce la regeneración parcial de la piel de mamíferos adultos". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 86 (3): 933–7. Bibcode :1989PNAS...86..933Y. doi : 10.1073/pnas.86.3.933 . JSTOR 33315. PMC 286593 . PMID 2915988.
- ^ O'Leary R, Ponnambalam S, Wood EJ (septiembre de 2003). "Muerte de células miofibroblásticas inducida por pioglitazona: implicaciones para la cicatrización cutánea". The British Journal of Dermatology . 149 (3): 665–7. doi :10.1046/j.1365-2133.2003.05501.x. PMID 14511015. S2CID 45852269.
- ^ Tong M, Tuk B, Hekking IM, Vermeij M, Barritault D, van Neck JW (2009). "Neovascularización estimulada, resolución de la inflamación y maduración del colágeno en la curación de heridas cutáneas de ratas mediante un mimético de glicosaminoglicano de heparán sulfato, OTR4120". Reparación y regeneración de heridas . 17 (6): 840–52. doi :10.1111/j.1524-475X.2009.00548.x. PMID 19903305. S2CID 17262546.
- ^ Barritault D, Caruelle JP (marzo de 2006). "[Agentes regeneradores (RGTAs): un nuevo enfoque terapéutico]" [Agentes regeneradores (RGTAs): un nuevo enfoque terapéutico]. Annales Pharmaceutiques Françaises (en francés). 64 (2): 135–44. doi :10.1016/S0003-4509(06)75306-8. PMID 16568015.
- ^ Van Neck et al, Los miméticos de proteoglicanos de sulfato de heparán mejoran la regeneración tisular: una descripción general. En el libro de Intech, con el título provisional "Regeneración tisular", ISBN 978-953-307-876-2 , se prevé su publicación en línea el 26 de noviembre de 2011" [ página necesaria ]
- ^ Personal de eurekalert.org (3 de junio de 2015). "Un científico del LIMR lidera un estudio que demuestra la regeneración tisular inducida por fármacos". eurekalert.org . Instituto Lankenau de Investigación Médica (LIMR). Archivado desde el original el 4 de julio de 2015 . Consultado el 3 de julio de 2015 .
- ^ Zhang Y, Strehin I, Bedelbaeva K, Gourevitch D, Clark L, Leferovich J, Messersmith PB, Heber-Katz E. Regeneración inducida por fármacos en ratones adultos. Sci Transl Med. 2015;290.
- ^ Rush, J. (2005). Tatuaje espiritual: una historia cultural del tatuaje, la perforación, la escarificación, el marcado y los implantes, Frog Ltd.
- ^ Brown BC, McKenna SP, Siddhi K, McGrouther DA, Bayat A (septiembre de 2008). "El costo oculto de las cicatrices en la piel: calidad de vida después de la cicatrización de la piel". Revista de cirugía plástica, reconstructiva y estética . 61 (9): 1049–58. doi :10.1016/j.bjps.2008.03.020. PMID 18617450.
- ^ Bayat A, McGrouther DA, Ferguson MW (enero de 2003). "Cicatrices en la piel". BMJ . 326 (7380): 88–92. doi :10.1136/bmj.326.7380.88. PMC 1125033 . PMID 12521975.
- ^ Clark, R. (1996). La biología molecular y celular de la reparación de heridas, Springer Us.
- ^ Tonnesen MG, Feng X, Clark RA (diciembre de 2000). "Angiogénesis en la cicatrización de heridas". The Journal of Investigative Dermatology. Actas del simposio . 5 (1): 40–6. doi : 10.1046/j.1087-0024.2000.00014.x . PMID 11147674.
- ^ ab Ferguson MW, Whitby DJ, Shah M, Armstrong J, Siebert JW, Longaker MT (abril de 1996). "Formación de cicatrices: la naturaleza espectral de la reparación de heridas fetales y adultas". Cirugía Plástica y Reconstructiva . 97 (4): 854–60. doi :10.1097/00006534-199604000-00029. PMID 8628785.
- ^ Brockes JP, Kumar A, Velloso CP (2001). "La regeneración como variable evolutiva". Journal of Anatomy . 199 (Pt 1–2): 3–11. doi :10.1046/j.1469-7580.2001.19910003.x. PMC 1594962 . PMID 11523827.
- ^ ab "Curación sin cicatrices". Star . Christchurch, Nueva Zelanda. 1906-07-07. pp. Página 4. Archivado desde el original el 2013-10-08 . Consultado el 2013-07-02 .
- ^ ab "Curación sin cicatrices". Marlborough Express, volumen XXXIX, número 160. paperspast.natlib.govt.nz. 12 de julio de 1906. pp. Página 1. Archivado desde el original el 8 de octubre de 2013. Consultado el 2 de julio de 2013 .
- ^ ab "Una maravillosa nueva cirugía". Reading Eagle . 1906-07-06. pp. Página 6. Archivado desde el original el 2016-03-12 . Consultado el 2013-07-02 .
- ^ Karin M, Clevers H (enero de 2016). "La inflamación reparadora se encarga de la regeneración tisular". Nature . 529 (7586): 307–15. Bibcode :2016Natur.529..307K. doi :10.1038/nature17039. PMC 5228603 . PMID 26791721.
- ^ Vlahopoulos SA (agosto de 2017). "El control aberrante de NF-κB en el cáncer permite la plasticidad transcripcional y fenotípica, para reducir la dependencia del tejido del huésped: modo molecular". Cancer Biology & Medicine . 14 (3): 254–270. doi :10.20892/j.issn.2095-3941.2017.0029. PMC 5570602 . PMID 28884042.
- ^ J Medicamentos Dermatol. 2019;18(1):9-16.
- ^ ab Zeng R, Lin C, Lin Z, Chen H, Lu W, Lin C, Li H (noviembre de 2018). "Enfoques para la cicatrización de heridas cutáneas: conceptos básicos y direcciones futuras". Investigación de células y tejidos . 374 (2): 217–232. doi :10.1007/s00441-018-2830-1. PMID 29637308. S2CID 4727434.
- ^ ab McLain NE, Moore ZE, Avsar P, et al. (Grupo Cochrane de Heridas) (marzo de 2021). "Limpieza de heridas para el tratamiento de las úlceras venosas de las piernas". Base de Datos Cochrane de Revisiones Sistemáticas . 2021 (3): CD011675. doi :10.1002/14651858.CD011675.pub2. PMC 8092712. PMID 33734426 .
- ^ Cumming BD, McElwain DL, Upton Z (enero de 2010). "Un modelo matemático de curación de heridas y cicatrización posterior". Journal of the Royal Society, Interface . 7 (42): 19–34. doi :10.1098/rsif.2008.0536. PMC 2839370 . PMID 19324672.
- ^ Gurtner GC, Werner S, Barrandon Y, Longaker MT (mayo de 2008). "Reparación y regeneración de heridas". Nature . 453 (7193): 314–21. Bibcode :2008Natur.453..314G. doi :10.1038/nature07039. PMID 18480812. S2CID 205213660.
- ^ Gurtner GC, Dauskardt RH, Wong VW, Bhatt KA, Wu K, Vial IN, Padois K, Korman JM, Longaker MT (agosto de 2011). "Mejora de la formación de cicatrices cutáneas mediante el control del entorno mecánico: estudios en animales grandes y de fase I". Anales de Cirugía . 254 (2): 217–25. doi :10.1097/SLA.0b013e318220b159. PMID 21606834. S2CID 21111114.
- ^ Kuhl E, Steinmann P (junio de 2004). "Modelado computacional de la curación: una aplicación del método de fuerza material". Biomecánica y modelado en mecanobiología . 2 (4): 187–203. doi :10.1007/s10237-003-0034-3. PMID 14872320. S2CID 8070456.
- ^ Rouillard AD, Holmes JW (septiembre de 2012). "Regulación mecánica de la migración de fibroblastos y remodelación del colágeno en la curación de infartos de miocardio". The Journal of Physiology . 590 (18): 4585–602. doi :10.1113/jphysiol.2012.229484. PMC 3477759 . PMID 22495588.
- ^ abcd Velnar T, Bailey T, Smrkolj V (1 de octubre de 2009). "El proceso de cicatrización de heridas: una descripción general de los mecanismos celulares y moleculares". Revista de investigación médica internacional . 37 (5): 1528–42. doi : 10.1177/147323000903700531 . PMID 19930861. S2CID 2496871.
- ^ ab Armitage J, Lockwood S (1 de octubre de 2011). "Incisiones en la piel y cierre de heridas". Surgery (Oxford) . Wound Management. 29 (10): 496–501. doi :10.1016/j.mpsur.2011.06.022.
- ^ Toon CD, Lusuku C, Ramamoorthy R, Davidson BR, Gurusamy KS, et al. (Grupo Cochrane de Heridas) (septiembre de 2015). "Retirada temprana versus tardía del apósito después del cierre primario de heridas quirúrgicas limpias y limpias-contaminadas". Base de Datos Cochrane de Revisiones Sistemáticas . 2015 (9): CD010259. doi :10.1002/14651858.CD010259.pub3. PMC 7087443. PMID 26331392 .
- ^ Norman G, Dumville JC, Mohapatra DP, Owens GL, Crosbie EJ (marzo de 2016). "Antibióticos y antisépticos para la cicatrización de heridas quirúrgicas por segunda intención". Base de Datos Cochrane de Revisiones Sistemáticas . 3 (4): CD011712. doi :10.1002/14651858.CD011712.pub2. PMC 6599835 . PMID 27021482.
- ^ Vermeulen H, Ubbink D, Goossens A, de Vos R, Legemate D, et al. (Grupo Cochrane de Heridas) (26 de enero de 2004). "Apósitos y agentes tópicos para la cicatrización de heridas quirúrgicas por segunda intención". Base de Datos Cochrane de Revisiones Sistemáticas . 2012 (2): CD003554. doi :10.1002/14651858.CD003554.pub2. PMC 8407283. PMID 15106207 .
- ^ Dumville JC, Owens GL, Crosbie EJ, Peinemann F, Liu Z, et al. (Grupo Cochrane de Heridas) (junio de 2015). "Terapia de presión negativa para el tratamiento de heridas quirúrgicas que cicatrizan por segunda intención". Base de datos Cochrane de revisiones sistemáticas (6): CD011278. doi :10.1002/14651858.CD011278.pub2. PMC 10960640. PMID 26042534 .
- ^ Tabla 3-1 en: Mitchell RS, Kumar V, Abbas AK, Nelson F (2007). Robbins Basic Pathology (8.ª ed.). Filadelfia: Saunders. ISBN 978-1-4160-2973-1.
- ^ Stejskalová A, Almquist BD (julio de 2017). "Uso de biomateriales para reconfigurar el proceso de reparación de heridas". Biomaterials Science . 5 (8): 1421–1434. doi :10.1039/c7bm00295e. PMC 5576529 . PMID 28692083.
- ^ Vyas KS, Vasconez HC. Cicatrización de heridas: productos biológicos, sustitutos de la piel, biomembranas y andamiajes Archivado el 13 de abril de 2017 en Wayback Machine . Atención sanitaria. 2014; 2(3):356-400.
{kind=link}
Enlaces externos
