Bacillus subtilis
 | Este artículo tiene varios problemas. Ayúdenos a mejorarlo o a discutir estos problemas en la página de discusión . ( Aprenda cómo y cuándo eliminar estos mensajes )
|


{kind=link}
{kind=link}
| Bacillus subtilis | |
|---|---|
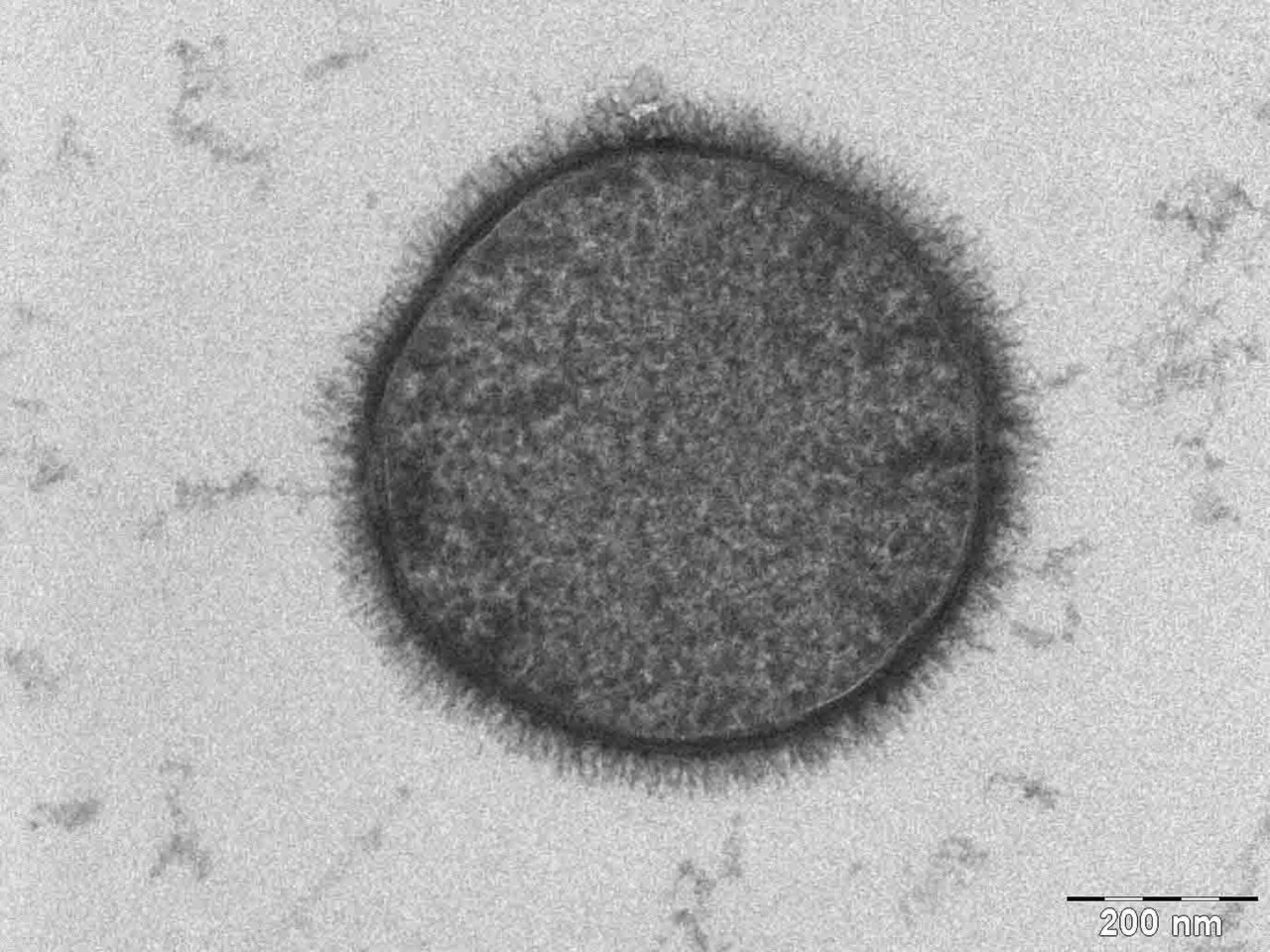 | |
| Micrografía TEM de una célula de B. subtilis en sección transversal (barra de escala = 200 nm ) | |
| Clasificación científica | |
| Dominio: | Bacteria |
| Filo: | Bacillota |
| Clase: | Bacilos |
| Orden: | Caryofanales |
| Familia: | Bacilláceas |
| Género: | Bacilo |
| Especies: | B. subtilis |
| Nombre binomial | |
| Bacillus subtilis ( Ehrenberg 1835) Cohn 1872 | |
| Sinónimos | |
| |
{kind=link}
Bacillus subtilis ( / b ə ˈ s ɪ l . ə s sub b ˈ t iː . l i s / ), [3] [ 4] conocido también como bacilo del heno o bacilo de la hierba , es una bacteria grampositiva , catalasa -positiva, que se encuentra en el suelo y el tracto gastrointestinal de rumiantes , humanos y esponjas marinas. [5] [6] [7] [8] Como miembro del género Bacillus , B. subtilis tiene forma de bastón y puede formar una endospora protectora resistente, lo que le permite tolerar condiciones ambientales extremas. B. subtilis ha sido históricamente clasificado como un aerobio obligado , aunque existe evidencia de que es un anaerobio facultativo . B. subtilis se considera la bacteria Gram-positiva mejor estudiada y un organismo modelo para estudiar la replicación cromosómica bacteriana y la diferenciación celular. Es uno de los campeones bacterianos en la producción de enzimas secretadas y es utilizado a escala industrial por empresas de biotecnología. [5] [6] [7]
Descripción
Bacillus subtilis es una bacteria Gram-positiva , con forma de bastón y catalasa -positiva. Originalmente fue nombrada Vibrio subtilis por Christian Gottfried Ehrenberg , [9] y renombrada Bacillus subtilis por Ferdinand Cohn en 1872 [10] (subtilis es la palabra latina para "fino, delgado, esbelto"). Las células de B. subtilis son típicamente con forma de bastón, y miden alrededor de 4-10 micrómetros (μm) de largo y 0,25-1,0 μm de diámetro, con un volumen celular de alrededor de 4,6 fL en fase estacionaria. [6] [11]
Al igual que otros miembros del género Bacillus , puede formar una endospora , para sobrevivir a condiciones ambientales extremas de temperatura y desecación. [12] B. subtilis es un anaerobio facultativo [6] [13] y había sido considerado como un aerobio obligado hasta 1998. B. subtilis está fuertemente flagelado , lo que le da la capacidad de moverse rápidamente en líquidos.
Se ha demostrado que B. subtilis es muy susceptible a la manipulación genética y se ha adoptado ampliamente como organismo modelo para estudios de laboratorio, especialmente de esporulación , que es un ejemplo simplificado de diferenciación celular . En términos de popularidad como organismo modelo de laboratorio, B. subtilis se considera a menudo como el equivalente grampositivo de Escherichia coli , una bacteria gramnegativa ampliamente estudiada. [14]
Características
Las características morfológicas, fisiológicas y bioquímicas de la colonia de Bacillus subtilis se muestran en la siguiente tabla. [6]
| Tipo de prueba | Prueba | Características |
|---|---|---|
| Personajes de la colonia | Tamaño | Medio |
| Tipo | Redondo | |
| Color | Blancuzco | |
| Forma | Convexo | |
| Caracteres morfológicos | Forma | Vara |
| Caracteres fisiológicos | Motilidad | + |
| Crecimiento al 6,5% de NaCl | + | |
| Caracteres bioquímicos | Tinción de Gram | + |
| Oxidasa | - | |
| Catalasa | + | |
| Oxidativo-fermentativo | Fermentativo | |
| Motilidad | - | |
| Rojo de metilo | - | |
| Proskauer de los Voges | + | |
| Indol | - | |
| Producción de H2S | + | |
| Ureasa | - | |
| Nitrato reductasa | + | |
| β-galactosidasa | + | |
| Hidrólisis de | Gelatina | + |
| Esculina | + | |
| Caseína | + | |
| Preadolescente de 40 años | + | |
| Preadolescente de 60 años | + | |
| Preadolescente de 80 años | + | |
| Producción de ácido a partir de | Glicerol | + |
| Galactosa | + | |
| D-glucosa | + | |
| D-fructosa | + | |
| D-manosa | + | |
| Manitol | + | |
| N-acetilglucosamina | + | |
| Amigdalina | + | |
| Maltosa | + | |
| D-Melibiosa | + | |
| D-trehalosa | + | |
| Glucógeno | + | |
| D-turanosa | + |
Nota: + = Positivo, – = Negativo
Hábitat
Esta especie se encuentra comúnmente en las capas superiores del suelo y se cree que B. subtilis es un comensal intestinal normal en los humanos. Un estudio de 2009 comparó la densidad de esporas encontradas en el suelo (alrededor de 10 6 esporas por gramo) con la encontrada en las heces humanas (alrededor de 10 4 esporas por gramo). La cantidad de esporas encontradas en el intestino humano fue demasiado alta para atribuirse únicamente al consumo a través de la contaminación de los alimentos. [15] En algunos hábitats de abejas, B. subtilis aparece en la flora intestinal de las abejas melíferas . [16] B. subtilis también se puede encontrar en entornos marinos. [6] [7]
Hay evidencia de que B. subtilis es saprofita por naturaleza. Los estudios han demostrado que la bacteria exhibe crecimiento vegetativo en suelo rico en materia orgánica y que las esporas se formaron cuando se agotaron los nutrientes. [17] Además, se ha demostrado que B. subtilis forma biopelículas en las raíces de las plantas, lo que podría explicar por qué se encuentra comúnmente en los microbiomas intestinales. [17] Tal vez los animales que comen plantas con biopelículas de B. subtilis pueden fomentar el crecimiento de la bacteria en su tracto gastrointestinal. Se ha demostrado que todo el ciclo de vida de B. subtilis puede completarse en el tracto gastrointestinal, lo que da credibilidad a la idea de que la bacteria ingresa al intestino a través del consumo de plantas y permanece presente como resultado de su capacidad para crecer en el intestino. [17]
Reproducción
Bacillus subtilis puede dividirse simétricamente para formar dos células hijas (fisión binaria), o asimétricamente, produciendo una única endospora que puede permanecer viable durante décadas y es resistente a condiciones ambientales desfavorables como sequía , salinidad , pH extremo , radiación y solventes . La endospora se forma en momentos de estrés nutricional y mediante el uso de hidrólisis, lo que permite que el organismo persista en el medio ambiente hasta que las condiciones se vuelvan favorables. Antes del proceso de esporulación, las células pueden volverse móviles produciendo flagelos , absorber ADN del medio ambiente o producir antibióticos . [6] [7] Estas respuestas se consideran intentos de buscar nutrientes buscando un entorno más favorable, lo que permite a la célula hacer uso de nuevo material genético beneficioso o simplemente eliminando la competencia. [ cita requerida ]
En condiciones de estrés, como la privación de nutrientes, B. subtilis experimenta el proceso de esporulación . Este proceso ha sido muy estudiado y ha servido como organismo modelo para estudiar la esporulación. [18]
Esporulación
Una vez que B. subtilis se compromete a esporular, se secreta el factor sigma sigma F. [20] Este factor promueve la esporulación. Se forma un septo de esporulación y un cromosoma se mueve lentamente hacia la preespora. Cuando un tercio de una copia de cromosoma está en la preespora y los dos tercios restantes están en la célula madre, el fragmento de cromosoma en la preespora contiene el locus para sigma F, que comienza a expresarse en la preespora. [21] Para prevenir la expresión de sigma F en la célula madre, se expresa un factor anti-sigma, que está codificado por spoIIAB, [22] . Cualquier factor anti-sigma residual en la preespora (que de otra manera interferiría con la esporulación) es inhibido por un factor anti-anti-sigma, que está codificado por spoIIAA. [22] SpoIIAA está ubicado cerca del locus para el factor sigma, por lo que se expresa constantemente en la preespora. Dado que el locus spoIIAB no se encuentra cerca de los loci sigma F y spoIIAA, se expresa solo en la célula madre y, por lo tanto, reprime la esporulación en esa célula, lo que permite que la esporulación continúe en la preespora. El spoIIAA residual en la célula madre reprime el spoIIAB, pero el spoIIAB se reemplaza constantemente, por lo que continúa inhibiendo la esporulación. Cuando el cromosoma completo se localiza en la preespora, el spoIIAB puede reprimir el sigma F. Por lo tanto, la asimetría genética del cromosoma de B. subtilis y la expresión de sigma F, spoIIAB y spoIIAA determinan la formación de esporas en B. subtilis.
{kind=link}

Replicación cromosómica
Bacillus subtilis es un organismo modelo utilizado para estudiar la replicación cromosómica bacteriana. La replicación del cromosoma circular único se inicia en un solo locus, el origen ( oriC ). La replicación procede de forma bidireccional y dos horquillas de replicación progresan en sentido horario y antihorario a lo largo del cromosoma. La replicación cromosómica se completa cuando las horquillas alcanzan la región terminal, que está situada opuesta al origen en el mapa cromosómico . La región terminal contiene varias secuencias cortas de ADN ( sitios Ter ) que promueven la detención de la replicación. Proteínas específicas median todos los pasos de la replicación del ADN. La comparación entre las proteínas implicadas en la replicación del ADN cromosómico en B. subtilis y en Escherichia coli revela similitudes y diferencias. Aunque los componentes básicos que promueven la iniciación, elongación y terminación de la replicación están bien conservados , se pueden encontrar algunas diferencias importantes (como que una bacteria carezca de proteínas esenciales en la otra). Estas diferencias subrayan la diversidad de los mecanismos y estrategias que han adoptado varias especies bacterianas para llevar a cabo la duplicación de sus genomas. [25]
Genoma
Bacillus subtilis tiene alrededor de 4.100 genes. De ellos, sólo 192 resultaron indispensables y se predijo que otros 79 también serían esenciales. Una gran mayoría de los genes esenciales se clasificaron en relativamente pocos dominios del metabolismo celular: aproximadamente la mitad participa en el procesamiento de la información, una quinta parte participa en la síntesis de la envoltura celular y la determinación de la forma y la división celular, y una décima parte está relacionada con la energía celular. [26]
La secuencia completa del genoma de la subcepa QB928 de B. subtilis tiene 4.146.839 pares de bases de ADN y 4.292 genes. La cepa QB928 se utiliza ampliamente en estudios genéticos debido a la presencia de varios marcadores [aroI(aroK)906 purE1 dal(alrA)1 trpC2]. [27]
En 2009 se caracterizaron varios ARN no codificantes en el genoma de B. subtilis , incluidos los ARN Bsr . [28] Los análisis genómicos comparativos basados en microarrays revelaron que los miembros de B. subtilis muestran una diversidad genómica considerable. [29]
FsrA es un ARN pequeño que se encuentra en Bacillus subtilis . Es un efector de la respuesta de ahorro de hierro y actúa para regular a la baja las proteínas que contienen hierro en momentos de baja biodisponibilidad de este. [30] [31]
Un probiótico prometedor para peces, la cepa WS1A de Bacillus subtilis , que posee actividad antimicrobiana contra Aeromonas veronii y suprimió la septicemia móvil por Aeromonas en Labeo rohita . El ensamblaje de novo resultó en un tamaño cromosómico estimado de 4.148.460 pb, con 4.288 marcos de lectura abiertos. [6] [7] El genoma de la cepa WS1A de B. subtilis contiene muchos genes potenciales, como los que codifican proteínas involucradas en la biosíntesis de riboflavina , vitamina B6 y aminoácidos ( ilvD ) y en la utilización del carbono ( pta ). [6] [7]
Transformación
La transformación bacteriana natural implica la transferencia de ADN de una bacteria a otra a través del medio circundante. En B. subtilis, la longitud del ADN transferido es mayor de 1271 kb (más de 1 millón de bases). [32] Es probable que el ADN transferido sea ADN bicatenario y, a menudo, represente más de un tercio de la longitud total del cromosoma de 4215 kb. [33] Parece que entre el 7 y el 9 % de las células receptoras ocupan un cromosoma entero. [34]
Para que una bacteria receptora se una, tome ADN exógeno de otra bacteria de la misma especie y lo recombine en su cromosoma, debe entrar en un estado fisiológico especial llamado competencia . La competencia en B. subtilis se induce hacia el final del crecimiento logarítmico, especialmente en condiciones de limitación de aminoácidos. [35] En estas condiciones estresantes de semiinanición, las células normalmente tienen solo una copia de su cromosoma y probablemente tengan un mayor daño en el ADN. Para probar si la transformación es una función adaptativa de B. subtilis para reparar su daño en el ADN, se realizaron experimentos utilizando luz ultravioleta como agente dañino. [36] [37] [38] Estos experimentos llevaron a la conclusión de que la competencia, con la absorción de ADN, es inducida específicamente por condiciones que dañan el ADN, y que la transformación funciona como un proceso para la reparación recombinatoria del daño en el ADN. [39]
Si bien el estado competente natural es común en los aislados de B. subtilis de laboratorio y de campo, algunas cepas industrialmente relevantes, por ejemplo, B. subtilis (natto), son reacias a la absorción de ADN debido a la presencia de sistemas de modificación de restricción que degradan el ADN exógeno. Los mutantes de B. subtilis (natto), que son defectuosos en una endonucleasa del sistema de modificación de restricción de tipo I, pueden actuar como receptores de plásmidos conjugativos en experimentos de apareamiento, allanando el camino para una mayor ingeniería genética de esta cepa particular de B. subtilis . [40]
Al adoptar la química verde en el uso de materiales menos peligrosos, a la vez que se ahorran costes, los investigadores han estado imitando los métodos de la naturaleza para sintetizar sustancias químicas que pueden ser útiles para la industria alimentaria y farmacéutica, "colocando moléculas a cuestas en cadenas cortas de ADN" antes de que se unan durante su apareamiento de bases complementarias entre las dos cadenas. Cada cadena llevará una molécula de interés particular que experimentará una reacción química específica simultáneamente cuando las dos cadenas correspondientes de pares de ADN se mantengan juntas como una cremallera, lo que permitirá que otra molécula de interés reaccione entre sí en una reacción controlada y aislada entre esas moléculas que se transportan en estas uniones complementarias de ADN. Al utilizar este método con ciertas bacterias que naturalmente siguen un proceso de replicación de múltiples pasos, los investigadores pueden llevar a cabo simultáneamente las interacciones de estas moléculas añadidas para interactuar con enzimas y otras moléculas utilizadas para una reacción secundaria tratándolas como una cápsula, que es similar a cómo las bacterias realizan sus propios procesos de replicación de ADN. [41]
Usos
Siglo XX
{kind=link}

Los cultivos de B. subtilis eran populares en todo el mundo, antes de la introducción de los antibióticos , como un agente inmunoestimulante para ayudar al tratamiento de enfermedades gastrointestinales y del tracto urinario . Se utilizó a lo largo de la década de 1950 como medicina alternativa , que tras la digestión se ha descubierto que estimula significativamente la actividad inmunitaria de amplio espectro, incluida la activación de la secreción de anticuerpos específicos IgM , IgG e IgA [42] y la liberación de dinucleótidos CpG que inducen la actividad productora de interferón IFN-α / IFNγ de leucocitos y citocinas importantes en el desarrollo de citotoxicidad hacia las células tumorales . [43] Se comercializó en toda América y Europa a partir de 1946 como una ayuda inmunoestimulante en el tratamiento de enfermedades intestinales y del tracto urinario como el rotavirus y la shigelosis . En 1966, el ejército de los EE. UU. arrojó Bacillus subtilis sobre las rejillas de las estaciones de metro de la ciudad de Nueva York durante cinco días para observar cómo un agente biológico distribuido alrededor de los trenes se dispersaría y potencialmente afectaría a los pasajeros desprevenidos. [44] Debido a su capacidad para sobrevivir, se cree que todavía está presente allí. [45]
El antibiótico bacitracina se aisló por primera vez de una variedad de Bacillus licheniformis llamada "Tracy I" [46] en 1945, que en ese entonces se consideraba parte de la especie B. subtilis . Todavía se fabrica comercialmente cultivando la variedad en un recipiente con un medio de cultivo líquido . Con el tiempo, la bacteria sintetiza bacitracina y secreta el antibiótico en el medio. Luego, la bacitracina se extrae del medio mediante procesos químicos. [47]
Desde la década de 1960, B. subtilis ha tenido una historia como especie de prueba en la experimentación de vuelos espaciales. Sus endosporas pueden sobrevivir hasta 6 años en el espacio si están recubiertas por partículas de polvo que las protegen de los rayos UV solares. [48] Se ha utilizado como un indicador de supervivencia extremófila en el espacio exterior , como en las misiones orbitales Exobiology Radiation Assembly , [49] [50] EXOSTACK , [51] [52] y EXPOSE . [53] [54] [55]
Los aislados naturales de tipo salvaje de B. subtilis son difíciles de manejar en comparación con las cepas de laboratorio que han sido sometidas a procesos de domesticación de mutagénesis y selección. Estas cepas a menudo tienen capacidades mejoradas de transformación (captación e integración de ADN ambiental), crecimiento y pérdida de capacidades necesarias "en la naturaleza". Y, aunque existen docenas de cepas diferentes que se ajustan a esta descripción, la cepa denominada '168' es la más utilizada. La cepa 168 es un auxótrofo del triptófano aislado después de la mutagénesis por rayos X de la cepa Marburg de B. subtilis y se utiliza ampliamente en investigación debido a su alta eficiencia de transformación. [56]
{kind=link}

Bacillus globigii , una especie estrechamente relacionada pero filogenéticamente distinta ahora conocida como Bacillus atrophaeus [57] [58] se utilizó como simulador de guerra biológica durante el Proyecto SHAD (también conocido como Proyecto 112 ). [59] El análisis genómico posterior mostró que las cepas utilizadas en esos estudios eran productos de un enriquecimiento deliberado de cepas que exhibían tasas anormalmente altas de esporulación . [60]
Una cepa de B. subtilis anteriormente conocida como Bacillus natto se utiliza en la producción comercial de la comida japonesa nattō , así como de la comida coreana similar cheonggukjang .
Siglo XXI
- Como organismo modelo, B. subtilis se utiliza comúnmente en estudios de laboratorio dirigidos a descubrir las propiedades y características fundamentales de las bacterias formadoras de esporas Gram-positivas. [29] En particular, los principios y mecanismos básicos que subyacen a la formación de la endospora duradera se han deducido de estudios de formación de esporas en B. subtilis .
- Sus propiedades de unión a la superficie desempeñan un papel en la eliminación segura de desechos de radionucleidos [por ejemplo, torio (IV) y plutonio (IV)]. [ cita requerida ]
- Debido a sus excelentes propiedades de fermentación, con altos rendimientos de producto (20 a 25 gramos por litro), se utiliza para producir diversas enzimas, como amilasa y proteasas. [61]
- B. subtilis se utiliza como inoculante de suelo en horticultura y agricultura . [62] [63] [64]
- Puede proporcionar algún beneficio a los productores de azafrán al acelerar el crecimiento del bulbo y aumentar el rendimiento de la biomasa del estigma. [65]
- Se utiliza como "organismo indicador" durante los procedimientos de esterilización por gas, para garantizar que un ciclo de esterilización se ha completado con éxito. En concreto, las endosporas de B. subtilis se utilizan para verificar que un ciclo ha alcanzado las condiciones de destrucción de esporas. [66] [67]
- Se ha descubierto que B. subtilis actúa como un fungicida bioproducto útil que previene el crecimiento de Monilinia vaccinii-corymbosi , también conocido como el hongo de la baya momia, sin interferir con la polinización o las cualidades de la fruta. [68]
- Se ha demostrado que las células de B. subtilis, tanto metabólicamente activas como no metabólicamente activas, reducen el oro (III) a oro (I) y oro (0) cuando hay oxígeno presente. Esta reducción biótica desempeña un papel en el ciclo del oro en sistemas geológicos y podría utilizarse potencialmente para recuperar oro sólido de dichos sistemas.
Subcepas nuevas y artificiales
- Se aislaron nuevas cepas de B. subtilis que podían utilizar 4-fluorotriptófano (4FTrp) pero no el triptófano canónico (Trp) para su propagación. Como el Trp solo está codificado por un único codón, hay evidencia de que el 4FTrp puede desplazarlo en el código genético. Los experimentos demostraron que el código genético canónico puede ser mutable. [69]
- Las cepas recombinantes pBE2C1 y pBE2C1AB se utilizaron en la producción de polihidroxialcanoatos (PHA), y los desechos de malta se pueden utilizar como fuente de carbono para una producción de PHA de menor costo. [ cita requerida ]
- Se utiliza para producir ácido hialurónico , que se emplea en el sector del cuidado de las articulaciones, en la asistencia sanitaria [70] y en la cosmética.
- Monsanto ha aislado un gen de B. subtilis que expresa la proteína de choque frío B y lo ha incorporado a su híbrido de maíz tolerante a la sequía MON 87460, cuya venta en los EE. UU. fue aprobada en noviembre de 2011. [71] [72]
- Se ha modificado una nueva cepa para convertir el néctar en miel mediante la secreción de enzimas. [73]
Seguridad
En otros animales
El Centro de Medicina Veterinaria de la FDA de EE. UU. revisó el Bacillus subtilis y encontró que no presenta problemas de seguridad cuando se usa en productos microbianos de alimentación directa, por lo que la Asociación de Funcionarios de Control de Alimentos de Estados Unidos lo incluyó aprobado para su uso como ingrediente de alimentos para animales en la Sección 36.14 "Microorganismos de alimentación directa". [ cita requerida ] La Sección de Alimentos para Producción y Salud Animal de la Agencia Canadiense de Inspección de Alimentos ha clasificado los ingredientes alimenticios aprobados deshidratados de cultivo de Bacillus como un aditivo de ensilaje bajo el Anexo IV-Parte 2-Clase 8.6 y le ha asignado el número internacional de ingrediente alimenticio IFN 8-19-119. [ cita requerida ] Por otro lado, la Autoridad Europea de Seguridad Alimentaria ha evaluado positivamente varios aditivos alimenticios que contienen esporas viables de B. subtilis , con respecto a su uso seguro para el aumento de peso en la producción animal.
En los humanos
Las esporas de Bacillus subtilis pueden sobrevivir al calor extremo generado durante la cocción. Algunas cepas de B. subtilis son responsables de causar la consistencia viscosa o el deterioro en forma de cuerda (una consistencia pegajosa y fibrosa causada por la producción bacteriana de polisacáridos de cadena larga ) en la masa de pan y los productos horneados en mal estado. [74] Durante mucho tiempo, la consistencia viscosa del pan se asoció únicamente con las especies de B. subtilis mediante pruebas bioquímicas. Los ensayos moleculares (ensayo de PCR de ADN polimórfico amplificado aleatoriamente, análisis de electroforesis en gel de gradiente desnaturalizante y secuenciación de la región V3 del ADN ribosómico 16S ) revelaron una mayor variedad de especies de Bacillus en panes viscosos, que parecen tener una actividad de amilasa positiva y una alta resistencia al calor. [75]
En un estudio de 16 semanas (10 días de administración de probióticos, seguidos de un período de lavado de 18 días por mes; se repitió el mismo procedimiento durante un total de 4 meses) se evaluó la B. subtilis CU1 (2 × 10 9 esporas por día) en sujetos sanos. Se descubrió que la B. subtilis CU1 era segura y bien tolerada por los sujetos sin efectos secundarios. [76]
Bacillus subtilis y sustancias derivadas de él han sido evaluadas por diferentes organismos autorizados para su uso seguro y beneficioso en alimentos. En los Estados Unidos, una carta de opinión emitida a principios de la década de 1960 por la Administración de Alimentos y Medicamentos (FDA) designó algunas sustancias derivadas de microorganismos como generalmente reconocidas como seguras (GRAS), incluidas las enzimas carbohidrasa y proteasa de B. subtilis . Las opiniones se basaron en el uso de cepas no patógenas y no toxicogénicas de los respectivos organismos y en el uso de buenas prácticas de fabricación actuales. [77] La FDA declaró que las enzimas derivadas de la cepa B. subtilis eran de uso común en alimentos antes del 1 de enero de 1958, y que las cepas no toxigénicas y no patógenas de B. subtilis están ampliamente disponibles y se han utilizado de manera segura en una variedad de aplicaciones alimentarias. Esto incluye el consumo de soja fermentada japonesa, en forma de natto , que se consume comúnmente en Japón, y contiene hasta 10 8 células viables por gramo. Las habas fermentadas son reconocidas por su contribución a una flora intestinal saludable y a la ingesta de vitamina K2 ; durante esta larga historia de uso generalizado, el natto no ha sido implicado en eventos adversos potencialmente atribuibles a la presencia de B. subtilis . [ cita requerida ] El producto natto y el natto B. subtilis como su componente principal son FOSHU (Alimentos para uso específico en la salud) aprobados por el Ministerio de Salud, Trabajo y Bienestar de Japón como efectivos para la preservación de la salud. [78]
La Autoridad Europea de Seguridad Alimentaria ha otorgado al Bacillus subtilis el estatus de "Presunción Cualificada de Seguridad" . [79]
Véase también
Referencias
- ^ Euzéby JP (2008). "Bacillus". Lista de nombres procariotas con posición en la nomenclatura . Consultado el 18 de noviembre de 2008 .
- ^ Ambrosiano N (30 de junio de 1999). "Las pruebas de biodetectores de laboratorio deben ser seguras y el público debe estar bien informado". Nota de prensa . Laboratorios Nacionales de Los Álamos. Archivado desde el original el 21 de septiembre de 2008. Consultado el 18 de noviembre de 2008 .
- ^ "Cómo se pronuncia bacillus". Diccionario Cambridge .
- ^ "subtilis". Wikcionario . 10 de marzo de 2023.
- ^ ab Errington J, Aart LT (mayo de 2020). "Perfil microbiano: Bacillus subtilis: organismo modelo para el desarrollo celular y caballo de batalla industrial". Microbiología . 166 (5): 425–427. doi : 10.1099/mic.0.000922 . PMC 7376258 . PMID 32391747.
- ^ abcdefghi Paul SI, Rahman MM, Salam MA, Khan MA, Islam MT (15 de diciembre de 2021). "Identificación de bacterias asociadas a esponjas marinas de la isla de San Martín de la Bahía de Bengala, con énfasis en la prevención de la septicemia por Aeromonas móviles en Labeo rohita". Acuicultura . 545 : 737156. doi :10.1016/j.aquaculture.2021.737156. ISSN 0044-8486.
- ^ abcdef Rahman MM, Paul SI, Akter T, Tay AC, Foysal MJ, Islam MT (septiembre de 2020). "Secuencia de todo el genoma de Bacillus subtilis WS1A, una cepa promisoria de pescado aislada de una esponja marina de la Bahía de Bengala". Anuncios de recursos de microbiología . 9 (39). doi :10.1128/mra.00641-20. PMC 7516141 . PMID 32972930.
- ^ Paul SI, Rahman MM (octubre de 2022). Gill SR (ed.). "Borrador de la secuencia del genoma de Bacillus subtilis YBS29, un posible probiótico para peces que previene la septicemia por Aeromonas móviles en Labeo rohita". Anuncios de recursos de microbiología . 11 (10): e0091522. doi :10.1128/mra.00915-22. PMC 9583808 . PMID 36154193.
- ^ Ehrenberg CG (1835). Physikalische Abhandlungen der Koeniglichen Akademie der Wissenschaften zu Berlin aus den Jahren 1833–1835 . págs. 145–336.
- ^ Cohn F (1872). "Untersuchungen über Bacterian". Beiträge zur Biologie der Pflanzen . vol. 1. págs. 127–224.
- ^ Yu AC, Loo JF, Yu S, Kong SK, Chan TF (enero de 2014). "Monitoreo del crecimiento bacteriano mediante detección de pulsos resistivos ajustables con una técnica basada en poros". Applied Microbiology and Biotechnology . 98 (2): 855–62. doi :10.1007/s00253-013-5377-9. PMID 24287933. S2CID 2956197.
- ^ Madigan M, Martinko J, eds. (2005). Brock Biología de microorganismos (11.ª ed.). Prentice Hall. ISBN 978-0-13-144329-7.[ página necesaria ]
- ^ Nakano MM, Zuber P (1998). "Crecimiento anaeróbico de un "aerobio estricto" ( Bacillus subtilis )". Revisión anual de microbiología . 52 (1): 165–90. doi :10.1146/annurev.micro.52.1.165. PMID 9891797.
- ^ Ruiz N, Silhavy TJ (septiembre de 2022). "Cómo Escherichia coli se convirtió en la bacteria insignia de la biología molecular". Revista de bacteriología . 204 (9): e0023022. doi :10.1128/jb.00230-22. PMC 9487582 . PMID 35916528. S2CID 251254431.
- ^ Hong HA, Khaneja R, Tam NM, Cazzato A, Tan S, Urdaci M, Brisson A, Gasbarrini A, Barnes I, Cutting SM (marzo de 2009). "Bacillus subtilis aislado del tracto gastrointestinal humano". Investigación en microbiología . 160 (2): 134–43. doi : 10.1016/j.resmic.2008.11.002 . PMID 19068230.
- ^ Sudhagar S, Reddy PR, Nagalakshmi G (abril de 2017). "Influencia de la elevación en la estructuración de las comunidades bacterianas intestinales de Apis cerana Fab" (PDF) . Revista de entomología y zoología . 5 (3): 434–440.
- ^ abc Tan IS, Ramamurthi KS (junio de 2014). "Formación de esporas en Bacillus subtilis: esporulación de Bacillus subtilis". Informes de microbiología ambiental . 6 (3): 212–225. doi :10.1111/1758-2229.12130. PMC 4078662 . PMID 24983526.
- ^ McKenney PT, Driks A, Eichenberger P (enero de 2013). "La endospora de Bacillus subtilis: ensamblaje y funciones de la capa multicapa". Nature Reviews. Microbiology . 11 (1): 33–44. doi : 10.1038/nrmicro2921 . PMC 9910062 . PMID 23202530. S2CID 205498395.
- ^ Stephens C (enero de 1998). "Esporulación bacteriana: ¿una cuestión de compromiso?". Current Biology . 8 (2): R45–R48. doi : 10.1016/S0960-9822(98)70031-4 . PMID 9427639. S2CID 14126998.
- ^ Earl AM, Losick R, Kolter R (junio de 2008). "Ecología y genómica de Bacillus subtilis". Tendencias en microbiología . 16 (6): 269–275. doi :10.1016/j.tim.2008.03.004. PMC 2819312 . PMID 18467096.
- ^ Higgins D, Dworkin J (enero de 2012). "Progreso reciente en la esporulación de Bacillus subtilis". FEMS Microbiology Reviews . 36 (1): 131–148. doi :10.1111/j.1574-6976.2011.00310.x. PMC 3237856 . PMID 22091839.
- ^ ab Slonczewski J, John Watkins Foster, Zinser ER. 2020. Microbiología: una ciencia en evolución. Nueva York: WW Norton & Company.
- ^ Piggot PJ, Coote JG (diciembre de 1976). "Aspectos genéticos de la formación de endosporas bacterianas". Bacteriological Reviews . 40 (4): 908–962. doi :10.1128/MMBR.40.4.908-962.1976. PMC 413989 . PMID 12736.
- ^ Jabbari S, Heap JT, King JR (enero de 2011). "Modelado matemático de la red de iniciación de la esporulación en Bacillus subtilis que revela el papel dual de la molécula señal de detección de quórum PhrA" (PDF) . Boletín de biología matemática . 73 (1): 181–211. doi :10.1007/s11538-010-9530-7. PMID 20238180. S2CID 9875633.
- ^ Noirot P (2007). "Replicación del cromosoma de Bacillus subtilis". En Graumann P (ed.).Bacillus : Biología celular y molecular . Caister Academic Press. ISBN 978-1-904455-12-7.[ página necesaria ]
- ^ Kobayashi K, Ehrlich SD, Albertini A, Amati G, Andersen KK, Arnaud M, et al. (abril de 2003). "Genes esenciales de Bacillus subtilis". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 100 (8): 4678–83. Bibcode :2003PNAS..100.4678K. doi : 10.1073/pnas.0730515100 . JSTOR 3144001. PMC 153615 . PMID 12682299.
- ^ Yu CS, Yim KY, Tsui SK, Chan TF (noviembre de 2012). "Secuencia completa del genoma de la cepa QB928 de Bacillus subtilis, una cepa ampliamente utilizada en estudios genéticos de B. subtilis". Journal of Bacteriology . 194 (22): 6308–9. doi :10.1128/JB.01533-12. PMC 3486399 . PMID 23105055.
- ^ Saito S, Kakeshita H, Nakamura K (enero de 2009). "Nuevos genes codificadores de ARN pequeño en las regiones intergénicas de Bacillus subtilis". Gene . 428 (1–2): 2–8. doi :10.1016/j.gene.2008.09.024. PMID 18948176.
- ^ ab Earl AM, Losick R, Kolter R (junio de 2008). "Ecología y genómica de Bacillus subtilis". Tendencias en microbiología . 16 (6): 269–75. doi :10.1016/j.tim.2008.03.004. PMC 2819312 . PMID 18467096.
- ^ Gaballa A, Antelmann H, Aguilar C, Khakh SK, Song KB, Smaldone GT, Helmann JD (agosto de 2008). "La respuesta de ahorro de hierro de Bacillus subtilis está mediada por un ARN pequeño regulado por Fur y tres proteínas pequeñas y básicas". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 105 (33): 11927–32. Bibcode :2008PNAS..10511927G. doi : 10.1073/pnas.0711752105 . PMC 2575260 . PMID 18697947.
- ^ Smaldone GT, Antelmann H, Gaballa A, Helmann JD (mayo de 2012). "El ARNm pequeño FsrA y la proteína FbpB median la inducción dependiente de hierro de las oxidasas que contienen hierro y azufre lutABC de Bacillus subtilis". Journal of Bacteriology . 194 (10): 2586–93. doi :10.1128/JB.05567-11. PMC 3347220 . PMID 22427629.
- ^ Saito Y, Taguchi H, Akamatsu T (marzo de 2006). "Destino del genoma bacteriano transformante tras su incorporación a células competentes de Bacillus subtilis: una longitud continua de ADN incorporado". Journal of Bioscience and Bioengineering . 101 (3): 257–62. doi :10.1263/jbb.101.257. PMID 16716928.
- ^ Saito Y, Taguchi H, Akamatsu T (abril de 2006). "El ADN introducido en las células competentes de Bacillus subtilis mediante la transformación de protoplastos lisados no es ADNmc sino ADNdc". Journal of Bioscience and Bioengineering . 101 (4): 334–39. doi :10.1263/jbb.101.334. PMID 16716942.
- ^ Akamatsu T, Taguchi H (abril de 2001). "Incorporación de todo el ADN cromosómico en lisados de protoplastos en células competentes de Bacillus subtilis". Biociencia, biotecnología y bioquímica . 65 (4): 823–29. doi : 10.1271/bbb.65.823 . PMID 11388459. S2CID 30118947.
- ^ Anagnostopoulos C, Spizizen J (mayo de 1961). "Requisitos para la transformación en Bacillus subtilis". Revista de bacteriología . 81 (5): 741–46. doi :10.1128/JB.81.5.741-746.1961. PMC 279084 . PMID 16561900.
- ^ Hoelzer MA, Michod RE (junio de 1991). "Reparación del ADN y evolución de la transformación en Bacillus subtilis. III. Sexo con ADN dañado". Genética . 128 (2): 215–23. doi :10.1093/genetics/128.2.215. PMC 1204460 . PMID 1906416.
- ^ Michod RE, Wojciechowski MF, Hoelzer MA (enero de 1988). "Reparación del ADN y evolución de la transformación en la bacteria Bacillus subtilis". Genética . 118 (1): 31–39. doi :10.1093/genética/118.1.31. PMC 1203263 . PMID 8608929.
- ^ Wojciechowski MF, Hoelzer MA, Michod RE (marzo de 1989). "Reparación del ADN y evolución de la transformación en Bacillus subtilis. II. Función de la reparación inducible". Genética . 121 (3): 411–22. doi :10.1093/genetics/121.3.411. PMC 1203629 . PMID 2497048.
- ^ Michod RE, Bernstein H, Nedelcu AM (mayo de 2008). "Valor adaptativo del sexo en patógenos microbianos". Infección, genética y evolución . 8 (3): 267–85. doi :10.1016/j.meegid.2008.01.002. PMID 18295550.
- ^ Itaya M, Nagasaku M, Shimada T, Ohtani N, Shiwa Y, Yoshikawa H, et al. (febrero de 2019). "Entrega estable y eficiente de ADN a Bacillus subtilis (natto) utilizando plásmidos de transferencia conjugacional pLS20". FEMS Microbiology Letters . 366 (4). doi :10.1093/femsle/fnz032. PMID 30726909.
- ^ "Química al natural: imitando los métodos limpios y eficientes de la naturaleza. - Biblioteca gratuita en línea". www.thefreelibrary.com . Consultado el 29 de abril de 2023 .
- ^ Ciprandi G, Scordamaglia A, Venuti D, Caria M, Canonica GW (diciembre de 1986). "Efectos in vitro de Bacillus subtilis sobre la respuesta inmune". Quimioterapia . 5 (6): 404–07. PMID 3100070.
- ^ Shylakhovenko VA (junio de 2003). "Efectos anticancerígenos e inmunoestimulantes de la fracción de nucleoproteína de 'Bacillus subtilis'"". Oncología experimental . 25 : 119–23.
- ^ Un estudio sobre la vulnerabilidad de los pasajeros del metro de la ciudad de Nueva York a la acción encubierta con agentes biológicos. Publicación miscelánea. Departamento del Ejército, Fort Detrick. 1968.
- ^ Rosoff S, Pontell H, Tillman R (2020). Lucro sin honor: delitos de cuello blanco y saqueo de Estados Unidos . Pearson. págs. 352-3. ISBN 9780134871486.
- ^ Podstawka A. "Bacillus licheniformis Tracy I | DSM 603, ATCC 10716, CCM 2181, IFO 12199, NBRC 12199, NCIB 8874, FDA BT1 | BacDiveID:686". bacdive.dsmz.de .
- ^ Johnson BA, Anker H, Meleney FL (octubre de 1945). "Bacitracina: un nuevo antibiótico producido por un miembro del grupo B. subtilis". Science . 102 (2650): 376–7. Bibcode :1945Sci...102..376J. doi :10.1126/science.102.2650.376. PMID 17770204. S2CID 51066.
- ^ Horneck G, Klaus DM, Mancinelli RL (marzo de 2010). "Microbiología espacial". Microbiology and Molecular Biology Reviews . 74 (1): 121–56. Bibcode :2010MMBR...74..121H. doi :10.1128/mmbr.00016-09. PMC 2832349 . PMID 20197502.
- ^ Dosis K, Bieger-Dosis A, Dillmann R, Gill M, Kerz O, Klein A, et al. (1995). "Experimento ERA "Bioquímica espacial"". Avances en la investigación espacial . 16 (8): 119–29. Bibcode :1995AdSpR..16h.119D. doi :10.1016/0273-1177(95)00280-R. PMID 11542696.
- ^ Vaisberg O, Fedorov A, Dunjushkin F, Kozhukhovsky A, Smirnov V, Avanov L, et al. (1995). "Poblaciones de iones en la cola de Venus". Avances en la investigación espacial . 16 (4): 105–18. Código Bibliográfico :1995AdSpR..16d.105V. doi :10.1016/0273-1177(95)00217-3.
- ^ Clancy P (23 de junio de 2005). Buscando vida, buscando el sistema solar . Cambridge University Press.[ página necesaria ]
- ^ Horneck G, Klaus DM, Mancinelli RL (marzo de 2010). "Microbiología espacial". Microbiology and Molecular Biology Reviews . 74 (1): 121–56. Bibcode :2010MMBR...74..121H. doi :10.1128/MMBR.00016-09. PMC 2832349 . PMID 20197502.
- ^ Fajardo-Cavazos P, Link L, Melosh HJ, Nicholson WL (diciembre de 2005). "Las esporas de Bacillus subtilis en meteoritos artificiales sobreviven a la entrada atmosférica a hipervelocidad: implicaciones para Lithopanspermia". Astrobiología . 5 (6): 726–36. Bibcode :2005AsBio...5..726F. doi :10.1089/ast.2005.5.726. PMID 16379527.
- ^ Brandstätter F, Brack A, Baglioni P, Cockell CS, Demets R, Edwards HG, et al. (2008). "Alteración mineralógica de meteoritos artificiales durante la entrada atmosférica. El experimento STONE-5". Ciencia planetaria y espacial . 56 (7): 976–84. Bibcode :2008P&SS...56..976B. CiteSeerX 10.1.1.549.4307 . doi :10.1016/j.pss.2007.12.014.
- ^ Wassmann M, Moeller R, Rabbow E, Panitz C, Horneck G, Reitz G, et al. (mayo de 2012). "Supervivencia de las esporas de la cepa MW01 de Bacillus subtilis resistente a los rayos UV tras la exposición a la órbita terrestre baja y a condiciones marcianas simuladas: datos del experimento espacial ADAPT en EXPOSE-E". Astrobiología . 12 (5): 498–507. Bibcode :2012AsBio..12..498W. doi :10.1089/ast.2011.0772. PMID 22680695.
- ^ Zeigler DR, Prágai Z, Rodriguez S, Chevreux B, Muffler A, Albert T, et al. (noviembre de 2008). "Los orígenes de 168, W23 y otras cepas heredadas de Bacillus subtilis". Revista de bacteriología . 190 (21): 6983–95. doi :10.1128/JB.00722-08. PMC 2580678 . PMID 18723616.
- ^ Nakamura LK (1989). "Relación taxonómica de las cepas de Bacillus subtilis pigmentadas de negro y una propuesta para Bacillus atrophaeus sp. nov". Revista internacional de bacteriología sistemática . 39 (3): 295–300. doi : 10.1099/00207713-39-3-295 .
- ^ Burke SA, Wright JD, Robinson MK, Bronk BV, Warren RL (mayo de 2004). "Detección de diversidad molecular en Bacillus atrophaeus mediante análisis de polimorfismo de longitud de fragmentos amplificados". Applied and Environmental Microbiology . 70 (5): 2786–90. Bibcode :2004ApEnM..70.2786B. doi :10.1128/AEM.70.5.2786-2790.2004. PMC 404429 . PMID 15128533.
- ^ "Proyecto 112/SHAD - Peligros y defensa a bordo". Departamento de Asuntos de Veteranos de los Estados Unidos. Archivado desde el original el 21 de febrero de 2015. Consultado el 25 de febrero de 2015 .
- ^ Gibbons HS, Broomall SM, McNew LA, Daligault H, Chapman C, Bruce D, Karavis M, Krepps M, McGregor PA, Hong C, Park KH, Akmal A, Feldman A, Lin JS, Chang WE, Higgs BW, Demirev P, Lindquist J, Liem A, Fochler E, Read TD, Tapia R, Johnson S, Bishop-Lilly KA, Detter C, Han C, Sozhamannan S, Rosenzweig CN, Skowronski EW (marzo de 2011). "Firmas genómicas de selección y mejora de cepas en Bacillus atrophaeus var. globigii, un simulador histórico de guerra biológica". PLOS ONE . 6 (3): e17836. Código Bibliográfico :2011PLoSO...617836G. doi : 10.1371/journal.pone.0017836 . PMC 3064580. PMID 21464989 .
- ^ van Dijl JM, Hecker M (enero de 2013). "Bacillus subtilis: de bacteria del suelo a fábrica de células supersecretoras". Factorías de células microbianas . 12 (3): 3. doi : 10.1186/1475-2859-12-3 . PMC 3564730 . PMID 23311580.
- ^ "Monilinia fructicola" (PDF) . Fichas técnicas sobre plagas de cuarentena . Fiscalía Europea (EPPO). Archivado desde el original (PDF) el 2015-06-04 . Consultado el 2015-07-21 .
- ^ Swain MR, Ray RC (2009). "Biocontrol y otras actividades beneficiosas de Bacillus subtilis aislado de la microflora del estiércol de vaca". Investigación microbiológica . 164 (2): 121–30. doi :10.1016/j.micres.2006.10.009. PMID 17320363.
- ^ Yánez-Mendizábal V (2011). "El control biológico de la podredumbre parda del melocotonero (Monilinia spp.) por Bacillus subtilis CPA-8 se basa en la producción de lipopéptidos similares a la fengicina". Revista Europea de Patología Vegetal . 132 (4): 609–19. doi :10.1007/s10658-011-9905-0. S2CID 15761522.
- ^ Sharaf-Eldin M, Elkholy S, Fernández JA, Junge H, Cheetham R, Guardiola J, Weathers P (agosto de 2008). "Bacillus subtilis FZB24 afecta la cantidad y calidad de las flores del azafrán (Crocus sativus)". Planta Medica . 74 (10): 1316–20. doi :10.1055/s-2008-1081293. PMC 3947403 . PMID 18622904.
- ^ "La Farmacopea Internacional – Cuarto Suplemento: Métodos de Análisis: 5. Procedimientos técnicos farmacéuticos: 5.8 Métodos de esterilización". Archivado desde el original el 8 de diciembre de 2008.
- ^ "AN-2203 Indicador biológico para EO (25/caja)". Andersen Products . Archivado desde el original el 2013-10-06 . Consultado el 2013-05-24 .
- ^ Ngugi HK, Dedej S, Delaplane KS, Savelle AT, Scherm H (1 de abril de 2005). "Efecto del biofungicida Serenade (Bacillus subtilis) aplicado a las flores sobre las variables relacionadas con la polinización en el arándano de ojo de conejo". Control biológico . 33 (1): 32–38. doi :10.1016/j.biocontrol.2005.01.002. ISSN 1049-9644.
- ^ Yu AC, Yim AK, Mat WK, Tong AH, Lok S, Xue H, Tsui SK, Wong JT, Chan TF (marzo de 2014). "Mutaciones que permiten el desplazamiento del triptófano por el 4-fluorotriptófano como aminoácido canónico del código genético". Genome Biology and Evolution . 6 (3): 629–41. doi :10.1093/gbe/evu044. PMC 3971595 . PMID 24572018.
- ^ "Preguntas frecuentes sobre hialuronato de sodio, preguntas frecuentes sobre ácido hialurónico, HA – Hyasis® | Novozymes Biopharma". Archivado desde el original el 28 de agosto de 2013. Consultado el 13 de agosto de 2013 .
- ^ Harrigan GG, Ridley WP, Miller KD, Sorbet R, Riordan SG, Nemeth MA, et al. (octubre de 2009). "El forraje y el grano de MON 87460, un híbrido de maíz tolerante a la sequía, son compositivamente equivalentes a los del maíz convencional". Journal of Agricultural and Food Chemistry . 57 (20): 9754–63. doi :10.1021/jf9021515. PMID 19778059.
- ^ USDA: Determinación del estado no regulado del maíz MON 87460 (Zea mays L)
- ^ Blum B (17 de noviembre de 2019). «Estudiantes israelíes ganan premio por elaborar miel sin abejas». Israel21c . Consultado el 24 de noviembre de 2019 .
- ^ "Descomposición de cuerdas | Procesos de horneado". BAKERpedia . 21 de junio de 2016 . Consultado el 7 de febrero de 2021 .
- ^ Pepe O, Blaiotta G, Moschetti G, Greco T, Villani F (abril de 2003). "Cepas productoras de cuerdas de Bacillus spp. a partir de pan de trigo y estrategia para su control por bacterias del ácido láctico". Microbiología Aplicada y Ambiental . 69 (4): 2321–9. Bibcode :2003ApEnM..69.2321P. doi :10.1128/AEM.69.4.2321-2329.2003. PMC 154770 . PMID 12676716.
- ^ Lefevre M, Racedo SM, Denayrolles M, Ripert G, Desfougères T, Lobach AR, et al. (febrero de 2017). "Evaluación de seguridad de Bacillus subtilis CU1 para su uso como probiótico en humanos". Toxicología y farmacología regulatorias . 83 : 54–65. doi : 10.1016/j.yrtph.2016.11.010 . PMID 27825987.
- ^ "Lista parcial de microorganismos de la FDA". Administración de Alimentos y Medicamentos. 2002.
- ^ Shortt C (septiembre de 2005). "Perspectivas sobre alimentos para usos específicos en la salud (FOSHU)". En Gibson GR (ed.). Boletín de Ciencia y Tecnología de los Alimentos: Alimentos Funcionales . Vol. 1. Lectura: IFIS Publishing. págs. 7-1. ISBN 978-0-86014-193-8.
- ^ Panel de la EFSA sobre peligros biológicos (BIOHAZ) (2010). «Opinión científica sobre el mantenimiento de la lista de microorganismos de QPS añadidos intencionadamente a alimentos o piensos (actualización de 2010)». Revista de la EFSA . 8 (12): 1944. doi : 10.2903/j.efsa.2010.1944 .
Enlaces externos
 Medios relacionados con Bacillus subtilis en Wikimedia Commons
Medios relacionados con Bacillus subtilis en Wikimedia Commons- SubtiWiki "Información actualizada de todos los genes de Bacillus subtilis "
- Evaluación final de riesgos de Bacillus subtilis en EPA.gov. Archivado desde el original el 9 de septiembre de 2015.
- Navegador del genoma de Bacillus subtilis
- Cepa tipo de Bacillus subtilis en BacDive, la base de metadatos de diversidad bacteriana
{kind=link}