Aminoácidos no proteinogénicos

En bioquímica , los aminoácidos no codificados o no proteinogénicos son distintos de los 22 aminoácidos proteinogénicos (21 en eucariotas [nota 1] ), que están codificados naturalmente en el genoma de los organismos para el ensamblaje de proteínas. Sin embargo, más de 140 aminoácidos no proteinogénicos se producen de forma natural en las proteínas y miles más pueden producirse en la naturaleza o sintetizarse en el laboratorio. [1] Los aminoácidos sintetizados químicamente pueden denominarse aminoácidos no naturales. Los aminoácidos no naturales pueden prepararse sintéticamente a partir de sus análogos nativos mediante modificaciones como la alquilación de aminas, la sustitución de la cadena lateral, la ciclización de la extensión del enlace estructural y los reemplazos isostéricos dentro de la cadena principal del aminoácido. [2] Muchos aminoácidos no proteinogénicos son importantes:
- intermediarios en la biosíntesis,
- en la formación postraduccional de proteínas,
- en un papel fisiológico (por ejemplo, componentes de las paredes celulares bacterianas , neurotransmisores y toxinas ),
- compuestos farmacológicos naturales o artificiales,
- presente en meteoritos o utilizado en experimentos prebióticos (como el experimento de Miller-Urey ),
- Podrían ser neurotransmisores importantes, como el ácido γ-aminobutírico , [3] y
- Puede desempeñar un papel crucial en la bioenergética celular, como la creatina . [4]
Definición por negación

Técnicamente, cualquier compuesto orgánico con un grupo funcional amina (–NH 2 ) y un ácido carboxílico (–COOH) es un aminoácido. Los aminoácidos proteinogénicos son un pequeño subconjunto de este grupo que poseen un átomo de carbono central (α- o 2-) portador de un grupo amino, un grupo carboxilo, una cadena lateral y una conformación α-hidrógeno levo , con excepción de la glicina , que es aquiral , y la prolina , cuyo grupo amina es una amina secundaria y, en consecuencia, se le denomina con frecuencia iminoácido por razones tradicionales, aunque no imino.
El código genético codifica 20 aminoácidos estándar para su incorporación a las proteínas durante la traducción . Sin embargo, hay dos aminoácidos proteinogénicos adicionales: selenocisteína y pirrolisina . Estos aminoácidos no estándar no tienen un codón dedicado, sino que se agregan en lugar de un codón de terminación cuando está presente una secuencia específica, codón UGA y elemento SECIS para selenocisteína, [5] secuencia descendente UAG PYLIS para pirrolisina. [6] Todos los demás aminoácidos se denominan "no proteinogénicos".
- Selenocisteína. Este aminoácido contiene un grupo selenol en su carbono β.
- Pirrolisina. Este aminoácido se forma uniendo al grupo ε-amino de la lisina un anillo de pirrolina carboxilado.


Existen varios grupos de aminoácidos: [7]
- 20 aminoácidos estándar
- 22 aminoácidos proteinogénicos
- Más de 80 aminoácidos creados abióticamente en altas concentraciones.
- Aproximadamente 900 se producen por vías naturales.
- Se han incorporado más de 118 aminoácidos modificados a la proteína.
Estos grupos se superponen, pero no son idénticos. Los 22 aminoácidos proteinogénicos son biosintetizados por organismos y algunos, pero no todos, también son abióticos (se encuentran en experimentos prebióticos y meteoritos). Algunos aminoácidos naturales, como la norleucina , se incorporan de forma incorrecta a las proteínas debido a la infidelidad del proceso de síntesis de proteínas. Muchos aminoácidos, como la ornitina , son intermediarios metabólicos producidos biosintéticamente, pero no incorporados a las proteínas. La modificación postraduccional de los residuos de aminoácidos en las proteínas conduce a la formación de muchos aminoácidos proteínicos, pero no proteinogénicos. Otros aminoácidos se encuentran únicamente en mezclas abióticas (por ejemplo, la α-metilnorvalina). Más de 30 aminoácidos no naturales se han insertado de forma traduccional en proteínas en sistemas diseñados, pero no son biosintéticos. [7]
Nomenclatura
Además del sistema de numeración IUPAC para diferenciar los distintos carbonos de una molécula orgánica, asignando secuencialmente un número a cada carbono, incluidos los que forman un grupo carboxílico, los carbonos a lo largo de la cadena lateral de los aminoácidos también se pueden etiquetar con letras griegas, donde el carbono α es el carbono quiral central que posee un grupo carboxilo, una cadena lateral y, en los α-aminoácidos, un grupo amino; el carbono en los grupos carboxílicos no se cuenta. [8] (En consecuencia, los nombres IUPAC de muchos α-aminoácidos no proteinogénicos comienzan con 2-amino- y terminan en -ácido ico ).
Natural noyo-α-aminoácidos
La mayoría de los aminoácidos naturales son α-aminoácidos en configuración L , pero existen algunas excepciones.
No alfa

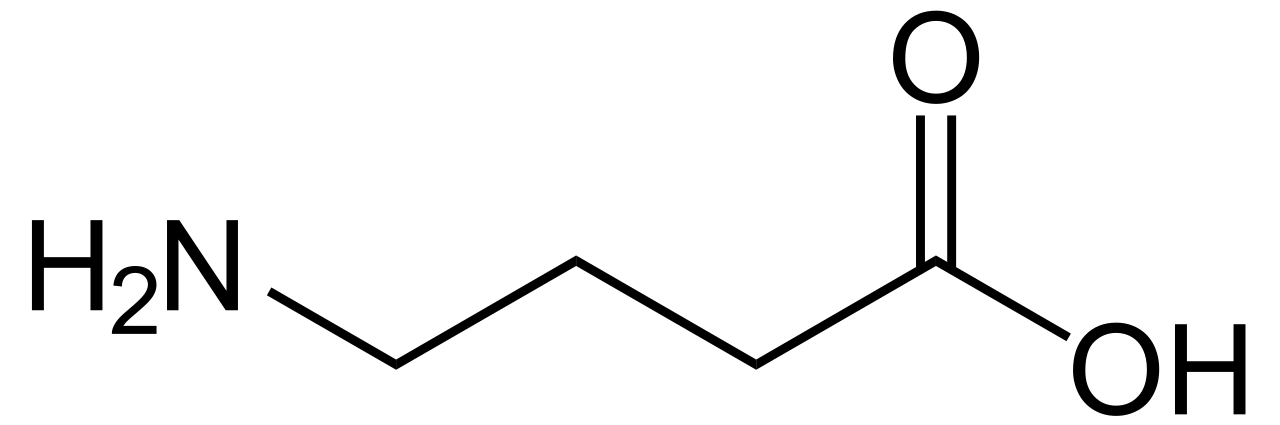
Existen algunos aminoácidos no alfa en los organismos. En estas estructuras, el grupo amino se desplaza más lejos del extremo del ácido carboxílico de la molécula de aminoácido. Por lo tanto, un β-aminoácido tiene el grupo amino unido al segundo carbono más alejado, y un γ-aminoácido lo tiene en el tercero. Algunos ejemplos son la β-alanina , el GABA y el ácido δ-aminolevulínico .
- β-alanina: un aminoácido producido por la aspartato 1-descarboxilasa y un precursor de la coenzima A [9] y los péptidos carnosina y anserina .
- Ácido γ-aminobutírico (GABA): un neurotransmisor en animales.
- Ácido δ-aminolevulínico: un intermediario en la biosíntesis de tetrapirrol ( hemo , clorofila , cobalamina , etc.).
- Ácido 4-aminobenzoico (PABA): un intermediario en la biosíntesis de folato
![β-alanina: un aminoácido producido por la aspartato 1-descarboxilasa y un precursor de la coenzima A[9] y los péptidos carnosina y anserina.](http://upload.wikimedia.org/wikipedia/commons/thumb/3/35/Beta-alanine_structure.svg/1280px-Beta-alanine_structure.svg.png)


El motivo por el que se utilizan los α-aminoácidos en las proteínas se ha relacionado con su frecuencia en meteoritos y experimentos prebióticos. [10] Una especulación inicial sobre las propiedades nocivas de los β-aminoácidos en términos de estructura secundaria [10] resultó ser incorrecta. [11]
D-aminoácidos
Algunos aminoácidos contienen la quiralidad absoluta opuesta, sustancias químicas que no están disponibles en la maquinaria normal de traducción y transcripción ribosómica. La mayoría de las paredes celulares bacterianas están formadas por peptidoglicano , un polímero compuesto de aminoazúcares reticulados con oligopéptidos cortos unidos entre sí. El oligopéptido no se sintetiza ribosómicamente y contiene varias peculiaridades, incluidos los D-aminoácidos , generalmente D -alanina y D -glutamato. Otra peculiaridad es que el primero es racemizado por enzimas de unión a PLP (codificadas por alr o el homólogo dadX ), mientras que el segundo es racemizado por una enzima independiente del cofactor ( murI ). Hay algunas variantes presentes, en Thermotoga spp. La D -lisina está presente y en ciertas bacterias resistentes a la vancomicina está presente la D -serina ( gen vanT ). [12] [13]
Sin un hidrógeno en el carbono α
Todos los aminoácidos proteinogénicos tienen al menos un hidrógeno en el carbono α. La glicina tiene dos hidrógenos y todos los demás tienen un hidrógeno y una cadena lateral. La sustitución del hidrógeno restante por un sustituyente más grande, como un grupo metilo, distorsiona la estructura de la proteína. [10]
En algunos hongos, el ácido α-aminoisobutírico se produce como precursor de péptidos, algunos de los cuales presentan propiedades antibióticas. [14] Este compuesto es similar a la alanina, pero posee un grupo metilo adicional en el carbono α en lugar de un hidrógeno. Por lo tanto, es aquiral. Otro compuesto similar a la alanina sin un hidrógeno α es la deshidroalanina , que posee una cadena lateral de metileno. Es uno de los varios deshidroaminoácidos naturales .
- alanina
- ácido aminoisobutírico
- deshidroalanina



Estereocentros de aminoácidos gemelos
Un subconjunto de L -α-aminoácidos son ambiguos en cuanto a cuál de los dos extremos es el carbono α. En las proteínas, un residuo de cisteína puede formar un enlace disulfuro con otro residuo de cisteína, reticulando así la proteína. Dos cisteínas reticuladas forman una molécula de cistina . La cisteína y la metionina se producen generalmente por sulfurilación directa, pero en algunas especies pueden producirse por transsulfuración , donde la homoserina o serina activada se fusiona a una cisteína u homocisteína formando cistationina . Un compuesto similar es la lantionina , que puede verse como dos moléculas de alanina unidas a través de un enlace tioéter y se encuentra en varios organismos. De manera similar, el ácido djenkolico , una toxina vegetal de las habas de jengkol , está compuesto de dos cisteínas conectadas por un grupo metileno. El ácido diaminopimélico se utiliza como puente en el peptidoglicano y se utiliza como precursor de la lisina (a través de su descarboxilación).
- cistina
- cistationina
- lantionina
- ácido djenkolico
- ácido diaminopimélico





Aminoácidos prebióticos y bioquímicas alternativas
En meteoritos y en experimentos prebióticos (por ejemplo, el experimento de Miller-Urey ) se encuentran muchos más aminoácidos que los veinte aminoácidos estándar, varios de los cuales están en concentraciones más altas que los estándares. Se ha conjeturado que si la vida basada en aminoácidos surgiera en otro lugar del universo, no más del 75% de los aminoácidos serían en común. [10] La anomalía más notable es la falta de ácido aminobutírico.
| Molécula | Descarga eléctrica | Meteorito de Murchinson |
|---|---|---|
| glicina | 100 | 100 |
| alanina | 180 | 36 |
| ácido α-amino- n -butírico | 61 | 19 |
| norvalina | 14 | 14 |
| Valina | 4.4 | |
| norleucina | 1.4 | |
| leucina | 2.6 | |
| isoleucina | 1.1 | |
| aloisoleucina | 1.2 | |
| t-leucina | < 0,005 | |
| ácido α-amino-n-heptanoico | 0.3 | |
| prolina | 0.3 | 22 |
| ácido pipecólico | 0,01 | 11 |
| Ácido α,β-diaminopropiónico | 1.5 | |
| Ácido α,γ-diaminobutírico | 7.6 | |
| ornitina | < 0,01 | |
| lisina | < 0,01 | |
| ácido aspártico | 7.7 | 13 |
| ácido glutámico | 1.7 | 20 |
| serina | 1.1 | |
| treonina | 0,2 | |
| alotreonina | 0,2 | |
| metionina | 0,1 | |
| homocisteína | 0,5 | |
| homoserina | 0,5 | |
| β-alanina | 4.3 | 10 |
| Ácido β-amino- n -butírico | 0,1 | 5 |
| ácido β-aminoisobutírico | 0,5 | 7 |
| ácido γ-aminobutírico | 0,5 | 7 |
| ácido α-aminoisobutírico | 7 | 33 |
| isovalina | 1 | 11 |
| sarcosina | 12.5 | 7 |
| N-etilglicina | 6.8 | 6 |
| N-propilglicina | 0,5 | |
| N-isopropilglicina | 0,5 | |
| N-metilalanina | 3.4 | 3 |
| N-etilalanina | < 0,05 | |
| N -metil-β-alanina | 1.0 | |
| N -etil-β-alanina | < 0,05 | |
| isoserina | 1.2 | |
| ácido α-hidroxi-γ-aminobutírico | 17 |
Cadena lateral recta
Se ha descrito el código genético como un accidente congelado y las razones por las que sólo hay un aminoácido estándar con una cadena lineal, la alanina, podrían ser simplemente la redundancia con valina, leucina e isoleucina. [10] Sin embargo, se informa que los aminoácidos de cadena lineal forman hélices alfa mucho más estables. [15]
- glicina (cadena lateral de hidrógeno)
- Alanina (cadena lateral de metilo)
- homoalanina o ácido α-aminobutírico (cadena lateral etílico)
- norvalina ( cadena lateral n -propil)
- norleucina ( cadena lateral n -butilo)
- homonorleucina ( cadena lateral n -Pentilo, ácido heptanoico mostrado)





Calcogen
La serina, homoserina , O -metilhomoserina y O -etilhomoserina poseen una cadena lateral de hidroximetilo, hidroxietilo, O -metilhidroximetilo y O -metilhidroxietilo; mientras que la cisteína, homocisteína , metionina y etionina poseen los equivalentes de tiol. Los equivalentes de selenol son selenocisteína, selenohomocisteína, selenometionina y selenoetionina. Los aminoácidos con el calcógeno inmediatamente inferior también se encuentran en la naturaleza: varias especies como Aspergillus fumigatus , Aspergillus terreus y Penicillium chrysogenum en ausencia de azufre son capaces de producir e incorporar a la proteína telurocisteína y telurometionina. [16]
Código genético ampliado
Roles
En las células, especialmente en las autótrofas, se encuentran varios aminoácidos no proteinogénicos como intermediarios metabólicos. Sin embargo, a pesar de la flexibilidad catalítica de las enzimas que se unen al PLP, muchos aminoácidos se sintetizan como cetoácidos (como el 4-metil-2-oxopentanoato a leucina) y se aminan en el último paso, lo que mantiene bastante bajo el número de intermediarios de aminoácidos no proteinogénicos.
La ornitina y la citrulina se encuentran en el ciclo de la urea , parte del catabolismo de los aminoácidos (véase más adelante). [17]
Además del metabolismo primario , varios aminoácidos no proteinogénicos son precursores o la producción final en el metabolismo secundario para formar pequeños compuestos o péptidos no ribosómicos (como algunas toxinas ).
Incorporado postraduccionalmente a la proteína
A pesar de no estar codificados por el código genético como aminoácidos proteinogénicos, algunos aminoácidos no estándar se encuentran sin embargo en las proteínas. Estos se forman por modificación postraduccional de las cadenas laterales de aminoácidos estándar presentes en la proteína diana. Estas modificaciones son a menudo esenciales para la función o regulación de una proteína; por ejemplo, en el γ-carboxiglutamato la carboxilación del glutamato permite una mejor unión de los cationes calcio , [18] y en la hidroxiprolina la hidroxilación de la prolina es crítica para el mantenimiento de los tejidos conectivos . [19] Otro ejemplo es la formación de hipusina en el factor de iniciación de la traducción EIF5A , a través de la modificación de un residuo de lisina. [20] Tales modificaciones también pueden determinar la localización de la proteína, por ejemplo, la adición de grupos hidrófobos largos puede hacer que una proteína se una a una membrana de fosfolípidos . [21]
- Ácido carboxiglutámico. Mientras que el ácido glutámico posee un grupo γ-carboxilo, el ácido carboxiglutámico posee dos.
- Hidroxiprolina. Este iminoácido se diferencia de la prolina debido a un grupo hidroxilo en el carbono 4.
- Hipusina . Este aminoácido se obtiene añadiendo al grupo ε-amino de una lisina una fracción 4-aminobutílica (obtenida a partir de la espermidina ).



-Pyroglutamic_acid_Structural_Formulae.png/1280px-(S)-Pyroglutamic_acid_Structural_Formulae.png)
Hay algunas evidencias preliminares de que el ácido aminomalónico puede estar presente, posiblemente por incorporación incorrecta, en las proteínas. [22] [23]
Análogos tóxicos
Varios aminoácidos no proteinogénicos son tóxicos debido a su capacidad de imitar ciertas propiedades de los aminoácidos proteinogénicos, como la tialisina . Algunos aminoácidos no proteinogénicos son neurotóxicos al imitar aminoácidos utilizados como neurotransmisores (es decir, no para la biosíntesis de proteínas), incluidos el ácido quiscuálico , la canavanina y el ácido azetidina-2-carboxílico . [24] La cefalosporina C tiene una cadena principal de ácido α-aminoadípico (homoglutamato) que está amidada con una fracción de cefalosporina. [25] La penicilamina es un aminoácido terapéutico, cuyo modo de acción se desconoce.
- Tialisina
- ácido quiscuálico
- canavanina
- ácido azetidina-2-carboxílico
- cefalosporina C
- penicilamina



-(-)-2-Azetidinecarboxylic_acid.svg/1280px-(S)-(-)-2-Azetidinecarboxylic_acid.svg.png)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
-Pyroglutamic_acid_Structural_Formulae.png){kind=link}
{kind=link}
{kind=link}
{kind=link}
-(-)-2-Azetidinecarboxylic_acid.svg){kind=link}
{kind=link}
{kind=link}
Las cianotoxinas naturales también pueden incluir aminoácidos no proteinogénicos. La microcistina y la nodularina , por ejemplo, son derivadas de ADDA , un β-aminoácido.
No aminoácidos
La taurina es un ácido aminosulfónico y no un ácido aminocarboxílico, aunque a veces se lo considera como tal ya que las cantidades necesarias para suprimir el auxótrofo en ciertos organismos (como los gatos) son más cercanas a las de los "aminoácidos esenciales" (auxotrofia de aminoácidos) que a las de las vitaminas (auxotrofia de cofactores).
Los osmolitos, sarcosina y glicina betaína se derivan de aminoácidos, pero tienen una amina secundaria y cuaternaria respectivamente.
Véase también
Notas
- ^ más formilmetionina en eucariotas con orgánulos procariotas como las mitocondrias
Referencias
- ^ Ambrogelly, A.; Palioura, S.; Söll, D. (2007). "Expansión natural del código genético". Nature Chemical Biology . 3 (1): 29–35. doi :10.1038/nchembio847. PMID 17173027.
- ^ Avan, Ilker; Hall, C. Dennis; Katritzky, Alan R. (22 de abril de 2014). "Peptidomiméticos a través de modificaciones de aminoácidos y enlaces peptídicos". Chemical Society Reviews . 43 (10): 3575–3594. doi :10.1039/C3CS60384A. PMID 24626261.
- ^ Sarasa, Sabna B.; Mahendran, Ramasamy; Muthusamy, Gayathri; Thankappan, Bency; Selta, Daniel Raja Femil; Angayarkanni, Jayaraman (2020). "Una breve revisión sobre el aminoácido no proteico, ácido gamma-aminobutírico (GABA): su producción y función en los microbios". Microbiología actual . 77 (4): 534–544. doi :10.1007/s00284-019-01839-w. PMID 31844936.
- ^ Ostojic, Sergej M. (1 de agosto de 2021). "Creatina como complemento alimenticio para la población general". Journal of Functional Foods . 83 : 104568. doi : 10.1016/j.jff.2021.104568 . ISSN 1756-4646.
- ^ Böck, A.; Forchhammer, K.; Heider, J.; Baron, C. (1991). "Síntesis de selenoproteína: una expansión del código genético". Tendencias en ciencias bioquímicas . 16 (12): 463–467. doi :10.1016/0968-0004(91)90180-4. PMID 1838215.
- ^ Théobald-Dietrich, A.; Giegé, R.; Rudinger-Thirion, JL (2005). "Evidencia de la existencia en los ARNm de un elemento de horquilla responsable de la inserción de pirrolisina dependiente del ribosoma en las proteínas". Biochimie . 87 (9–10): 813–817. doi :10.1016/j.biochi.2005.03.006. PMID 16164991.
- ^ ab Lu, Y.; Freeland, S. (2006). "Sobre la evolución del alfabeto estándar de aminoácidos". Genome Biology . 7 (1): 102. doi : 10.1186/gb-2006-7-1-102 . PMC 1431706 . PMID 16515719.
- ^ Voet, D.; Voet, JG (2004). Bioquímica (3ª ed.). John Wiley e hijos. ISBN 978-0471193500.
- ^ Chakauya, E.; Coxon, KM; Ottenhof, HH; Whitney, HM; Blundell, TL; Abell, C.; Smith, AG (2005). "Biosíntesis de pantotenato en plantas superiores". Biochemical Society Transactions . 33 (4): 743–746. doi :10.1042/BST0330743. PMID 16042590.
- ^ abcde Weber, AL; Miller, SL (1981). "Razones para la aparición de los veinte aminoácidos proteicos codificados". Journal of Molecular Evolution . 17 (5): 273–284. Bibcode :1981JMolE..17..273W. doi :10.1007/BF01795749. PMID 7277510. S2CID 27957755.
- ^ Koyack, MJ; Cheng, RP (2006). "Diseño y síntesis de péptidos β con actividad biológica". Diseño de proteínas . Métodos en biología molecular. Vol. 340. págs. 95-109. doi :10.1385/1-59745-116-9:95. ISBN. 978-1-59745-116-1. Número de identificación personal 16957334.
- ^ Boniface, A.; Parquet, C.; Arthur, M.; Mengin-Lecreulx, D.; Blanot, D. (2009). "La elucidación de la estructura del peptidoglicano de Thermotoga maritima revela dos nuevos tipos de enlaces cruzados". Journal of Biological Chemistry . 284 (33): 21856–21862. doi : 10.1074/jbc.M109.034363 . PMC 2755910 . PMID 19542229.
- ^ Arias, CA; Martín-Martínez, M.; Blundell, TL; Arthur, M.; Courvalin, P.; Reynolds, PE (1999). "Caracterización y modelado de vanT: una nueva serina racemasa unida a la membrana de Enterococcus gallinarum BM4174 resistente a la vancomicina". Microbiología molecular . 31 (6): 1653–1664. doi : 10.1046/j.1365-2958.1999.01294.x . PMID 10209740. S2CID 25796469.
- ^ Gao, X.; Chooi, YH; Ames, BD; Wang, P.; Walsh, CT; Tang, Y. (2011). "Biosíntesis de alcaloides indólicos fúngicos: investigación genética y bioquímica de la vía de la triptoquialanina en Penicillium aethiopicum". Revista de la Sociedad Química Estadounidense . 133 (8): 2729–2741. doi :10.1021/ja1101085. PMC 3045477 . PMID 21299212.
- ^ Padmanabhan, S.; Baldwin, RL (1991). "Los aminoácidos no polares de cadena recta son buenos formadores de hélice en agua". Journal of Molecular Biology . 219 (2): 135–137. doi : 10.1016/0022-2836(91)90553-I . PMID 2038048.
- ^ Ramadan, SE; Razak, AA; Ragab, AM; El-Meleigy, M. (1989). "Incorporación de telurio en aminoácidos y proteínas en hongos tolerantes al telurio". Biological Trace Element Research . 20 (3): 225–232. doi :10.1007/BF02917437. PMID 2484755. S2CID 9439946.
- ^ Curis, E.; Nicolis, I.; Moinard, C.; Osowska, S.; Zerrouk, N.; Bénazeth, S.; Cynober, L. (2005). "Casi todo sobre la citrulina en mamíferos". Aminoácidos . 29 (3): 177–205. doi :10.1007/s00726-005-0235-4. PMID 16082501. S2CID 23877884.
- ^ Vermeer, C. (1990). "Proteínas que contienen gamma-carboxiglutamato y la carboxilasa dependiente de la vitamina K". The Biochemical Journal . 266 (3): 625–636. doi :10.1042/bj2660625. PMC 1131186 . PMID 2183788.
- ^ Bhattacharjee, A; Bansal, M (2005). "Estructura del colágeno: la triple hélice de Madrás y el escenario actual". IUBMB Life . 57 (3): 161–172. doi : 10.1080/15216540500090710 . PMID 16036578. S2CID 7211864.
- ^ Park, MH (2006). "La síntesis postraduccional de un aminoácido derivado de poliamina, la hipusina, en el factor de iniciación de la traducción eucariota 5A (eIF5A)". Journal of Biochemistry . 139 (2): 161–169. doi :10.1093/jb/mvj034. PMC 2494880 . PMID 16452303.
- ^ Blenis, J; Resh, MD (1993). "Localización subcelular especificada por la acilación y fosforilación de proteínas". Current Opinion in Cell Biology . 5 (6): 984–989. doi :10.1016/0955-0674(93)90081-z. PMID 8129952.
- ^ Copley, SD; Frank, E.; Kirsch, WM; Koch, TH (1992). "Detección y posibles orígenes del ácido aminomalónico en hidrolizados proteicos". Analytical Biochemistry . 201 (1): 152–157. doi :10.1016/0003-2697(92)90188-D. PMID 1621954.
- ^ Van Buskirk, JJ; Kirsch, WM; Kleyer, DL; Barkley, RM; Koch, TH (1984). "Ácido aminomalónico: identificación en Escherichia coli y placa aterosclerótica". Actas de la Academia Nacional de Ciencias . 81 (3): 722–725. Bibcode :1984PNAS...81..722V. doi : 10.1073/pnas.81.3.722 . PMC 344907 . PMID 6366787.
- ^ Dasuri, K.; Ebenezer, PJ; Uranga, RM; Gavilán, E.; Zhang, L.; Fernandez-Kim, SOK; Bruce-Keller, AJ; Keller, JN (2011). "Toxicidad de análogos de aminoácidos en cultivos primarios de neuronas y astrocitos de rata: implicaciones para el plegamiento incorrecto de proteínas y la regulación de TDP-43". Journal of Neuroscience Research . 89 (9): 1471–1477. doi :10.1002/jnr.22677. PMC 3175609 . PMID 21608013.
- ^ Trown, PW; Smith, B.; Abraham, EP (1963). "Biosíntesis de cefalosporina C a partir de aminoácidos". The Biochemical Journal . 86 (2): 284–291. doi :10.1042/bj0860284. PMC 1201751 . PMID 13994319.
