Megaherbívoro
_(16485955207).jpg/1280px-Hippo_(Hippopotamus_amphibius)_(16485955207).jpg)
Los megaherbívoros (del griego μέγας megas "grande" y del latín herbivora "herbívoro" [1] ) son grandes herbívoros que pueden superar los 1000 kg (2200 lb) de peso. Los primeros herbívoros en alcanzar tales tamaños, como los parieasaurios, aparecieron en el período Pérmico . Durante la mayor parte del Mesozoico , el nicho de los megaherbívoros estuvo dominado en gran medida por los dinosaurios hasta su extinción durante el evento de extinción masiva del Cretácico-Paleógeno . Después de este período, las especies de pequeños mamíferos evolucionaron a grandes herbívoros en el Paleógeno . Como parte de las extinciones de la megafauna del Pleistoceno tardío , el 80% de las especies de megaherbívoros se extinguieron, y los megaherbívoros se extinguieron por completo en Europa, Australia y América. Los megaherbívoros recientes incluyen elefantes , rinocerontes , hipopótamos y jirafas . Existen nueve especies actuales de megaherbívoros terrestres que viven en África y Asia . El elefante africano de sabana es la especie más grande que existe.
Los megaherbívoros actuales son especies clave en su entorno. Deshojaron el paisaje y esparcieron una mayor cantidad de semillas que otros frugívoros . Los megaherbívoros actuales, como la mayoría de los grandes mamíferos, son especies seleccionadas por K. Se caracterizan por su gran tamaño, inmunidad relativa a la depredación , su efecto sobre las especies vegetales y su tolerancia dietética.
Definición
Los megaherbívoros son herbívoros grandes que pesan más de una tonelada cuando están completamente desarrollados. [2] Incluyen tanto herbívoros marinos como terrestres. [1] Son un tipo de megafauna (>45 kg) y son los animales más grandes de la tierra. [3]
Evolución
Pérmico
Los megaherbívoros evolucionaron por primera vez a principios del Pérmico (hace 300 millones de años). [4] Los primeros megaherbívoros fueron sinápsidos ; se volvieron algo raros después del evento de extinción masiva del Pérmico-Triásico . [5] [6] Los taxones consistían principalmente en dicinodontos , [7] y pareiasaurios . [8] La causa exacta de la extinción sigue siendo desconocida. Se cree que la principal causa de la extinción fueron las erupciones volcánicas de basalto de inundación que crearon las Trampas Siberianas , [9] que liberaron dióxido de azufre y dióxido de carbono , lo que resultó en euxinia , [10] elevando las temperaturas globales, [11] y acidificando los océanos . [12]
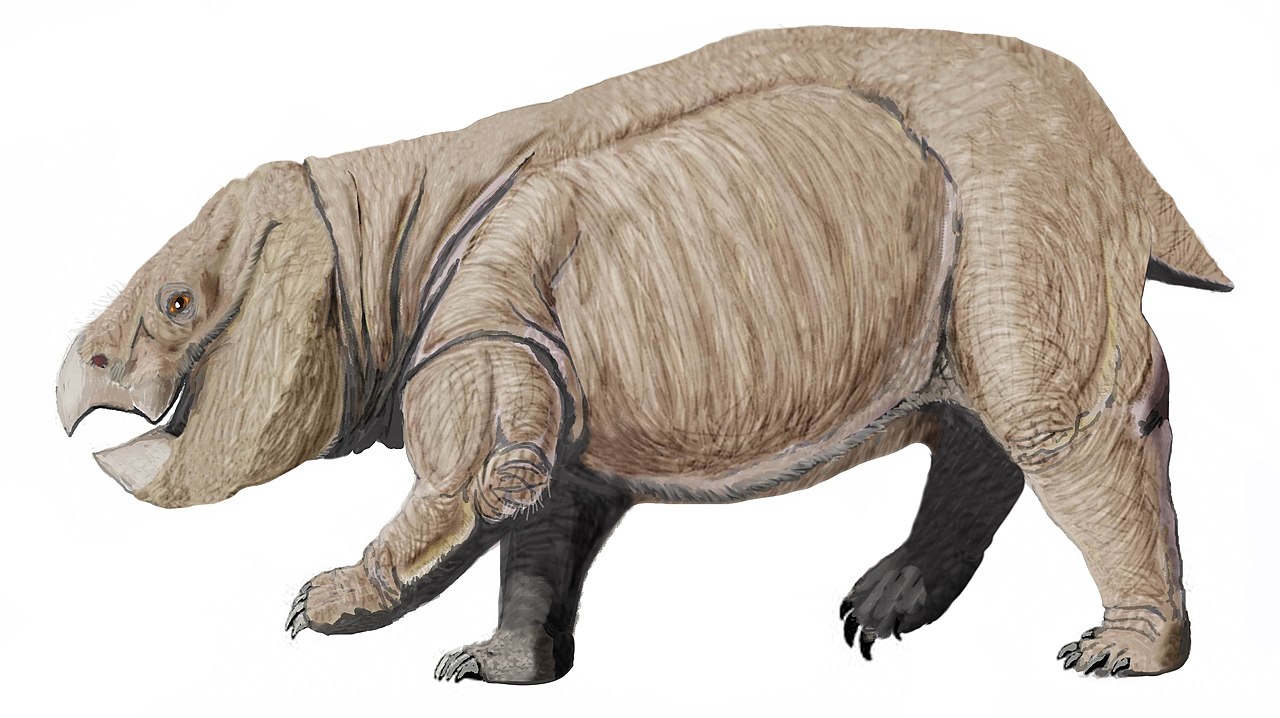
Triásico
Lisowicia fue el último dicinodonte que vivió y se extinguió en el Triásico Tardío. [5] Algunos científicos han propuesto que nunca hubo un evento de extinción del Triásico-Jurásico , pero otros sostienen que las extinciones ocurrieron antes. Sin embargo, se cree que los basaltos de inundación son el principal impulsor de las extinciones hacia el final del Triásico . [13] [14]
| Amniota |
| ||||||||||||||||||

jurásico
La estructura taxonómica luego cambió a sauropodomorfos . Otros taxones incluyeron estegosaurios y anquilosaurios . [15] El cambio en la taxonomía ocurrió aproximadamente al mismo tiempo con la divergencia de la vegetación predominante y con las extinciones . Los nuevos taxones pueden haber causado exclusión competitiva (es decir, predominando y eliminando a otros taxones), o pueden haber adoptado el nicho ecológico de grupos extintos. [4] [16]
Cretáceo







Desde el Triásico hasta el Cretácico , un conjunto diverso de dinosaurios megaherbívoros , como los saurópodos , [17] ocuparon diferentes nichos ecológicos. Según su dentición, los anquilosaurios pueden haber consumido principalmente plantas suculentas, a diferencia de los nodosaurios, que eran principalmente ramoneadores. Se cree que los ceratópsidos se alimentaban de vegetación accidentada, debido a que su mandíbula estaba diseñada para un efecto aplastante. Los estudios sobre la dentición de los hadrosaurios concluyeron que se alimentaban principalmente de frutas. [18]
Paleógeno
.jpg/1280px-%D0%9F%D0%B0%D0%BB%D0%B5%D0%BE%D0%BD%D1%82%D0%BE%D0%BB%D0%BE%D0%B3%D0%B8%D1%87%D0%B5%D1%81%D0%BA%D0%B8%D0%B9_%D0%BC%D1%83%D0%B7%D0%B5%D0%B9_%D0%9E%D1%80%D0%BB%D0%BE%D0%B2%D0%B0_(20221008151051).jpg)
.
Después de la extinción masiva del Cretácico-Paleógeno , los dinosaurios megaherbívoros fueron extirpados de la faz de la tierra. Se cree que un mecanismo jugó un papel importante: un evento de impacto extraterrestre en la península de Yucatán . [19] Durante aproximadamente 25 millones de años, la Tierra estuvo vacía de grandes herbívoros terrestres que pesaran más de 1 tonelada. Después de este período, las especies de pequeños mamíferos evolucionaron a grandes herbívoros. Los mamíferos herbívoros habían evolucionado al tamaño de megaherbívoros en todos los continentes alrededor de 40 millones de años atrás. [2] Los más grandes de estos animales fueron Paraceratheriidae y Proboscidea . [20] Otros taxones incluyeron Brontotheriidae . [21] Los Sirenia , megaherbívoros acuáticos, como Dugongidae , Protosirenidae y Prorastomidae estuvieron presentes en el Eoceno . [22] Los megaherbívoros habitaron todas las masas continentales importantes en el Cenozoico y el Pleistoceno antes de la llegada de los humanos . [4]
pleistoceno
Había alrededor de 50 especies diferentes en el Pleistoceno tardío : [3]
| Mamíferos |
| ||||||||||||||||||||||||||||||||||||||||||||||||
.jpg/1280px-Diprotodon_optatum_(2).jpg)




.jpg/1280px-Macrauchenia_(trunkless).jpg)


.jpg/1280px-Voyage_en_Abyssinie_Plate_2_(white_background).jpg)
.png/1280px-Cladogram_of_Cetacea_within_Artiodactyla_(Camelus_bactrianus).png)
_(14562088237)_white_background.jpg/1280px-Birds_and_nature_(1901)_(14562088237)_white_background.jpg)
Diprotodon , el marsupial más grande que jamás haya existido, estaba presente en todo el continente australiano a finales del Pleistoceno. [23] Además, los gliptodontes, como muchos otros xenartros, eran herbívoros de pastoreo, que no poseían dientes incisivos o caninos, pero tenían muelas que habrían podido triturar vegetación dura. [24] Los perezosos terrestres eran herbívoros, algunos eran ramoneadores , [25] otros pastaban , [26] y algunos eran intermedios entre los dos como alimentadores mixtos. [27] Los mamuts, como los elefantes modernos, tenían molares hipsodontes . Estas características les permitieron a los mamuts vivir una vida expansiva debido a la disponibilidad de pastos y árboles. [28] Hoy, nueve de las 50 especies persisten. Las Américas vieron la peor disminución de megaherbívoros, con las 27 especies extintas. [3]
La extinción masiva del Cuaternario es un evento en el que muchas especies de megafauna (en particular, mamíferos) se extinguieron. Este evento provocó la desaparición de megaherbívoros en la mayoría de los continentes de la Tierra. [29] El cambio climático y la llegada de los humanos se consideran causas probables de las extinciones. [30] Se cree que los humanos cazaron megaherbívoros hasta la extinción, lo que luego llevó a la extinción de los carnívoros y carroñeros que se habían aprovechado de esos animales. [31] [32] [33] Los científicos han propuesto que el clima cada vez más extremo (veranos más cálidos e inviernos más fríos), conocido como " continentalidad ", o los cambios relacionados en las precipitaciones causaron las extinciones. [34]
Reciente
| Placenta |
| ||||||||||||||||||||||||||||||


_(16485955207).jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
.png){kind=link}
_(14562088237)_white_background.jpg){kind=link}
{kind=link}
{kind=link}
Hay nueve especies existentes de megaherbívoros, que se encuentran en África y Asia. [35] [36] Incluyen elefantes, rinocerontes, hipopótamos y jirafas. [35] [37] : 1 [38] Los elefantes pertenecen al orden Proboscidea , que existe desde finales del Paleoceno . [39] Los hipopótamos son los parientes vivos más cercanos a los cetáceos . Poco después de que el ancestro común de las ballenas y los hipopótamos divergiera de los ungulados de dedos pares , los linajes de cetáceos e hipopótamos se separaron. [40] [41] Giraffidae es un taxón hermano de Antilocapridae , con una división estimada de hace más de 20 millones de años, según un estudio del genoma de 2019. [42] Los rinocerontes pueden tener su origen en Hyrachyus , un animal cuyos restos datan de finales del Eoceno . [37] : 17
Los megaherbívoros y otros grandes herbívoros son cada vez menos comunes en toda su distribución natural, lo que está teniendo un impacto en las especies animales dentro del ecosistema. Esto se atribuye principalmente a la destrucción de su entorno natural , la agricultura , la caza excesiva y la invasión humana de sus hábitats. [43] [44] Como consecuencia de su lenta tasa de reproducción y la preferencia por apuntar a especies más grandes, la sobreexplotación plantea la mayor amenaza para los megaherbívoros. A medida que pasa el tiempo, se cree que la situación solo empeorará. [43]
Ecología de la megaherbivoría reciente
Navegadores y pastores
Las especies vivas presentan las siguientes adaptaciones: tienen tolerancia dietética, un fuerte efecto sobre la vegetación y, con excepción de las crías, enfrentan pocas amenazas de los depredadores. [45] [46] [47]
Los elefantes y los rinocerontes indios exhiben hábitos alimentarios tanto de pastoreo como de ramoneo. El hipopótamo y el rinoceronte blanco prefieren la herbivoría de pastoreo , mientras que las jirafas y las otras tres especies de rinocerontes seleccionan con mayor frecuencia la herbivoría de ramoneo . [48] Los megaherbívoros mamíferos consumen predominantemente gramíneas . Prefieren comer las hojas y el tallo de la planta , así como sus frutos . También exhiben fermentación tanto del intestino anterior como del intestino posterior , con los rinocerontes, hipopótamos y elefantes mostrando la primera y las jirafas mostrando la segunda. [37] : 16 Su tasa metabólica es letárgica y, como resultado, la digestión se ralentiza. Durante este período de digestión prolongado, la materia vegetal rica en fibra se desintegra. [38]
Debido a su tamaño, los megaherbívoros pueden defoliar el paisaje; por ello, se consideran especies clave en su entorno. [49] Los megaherbívoros afectan la composición de las especies vegetales, lo que altera el movimiento y el intercambio de materia inorgánica y orgánica para la producción de materia. Pueden abrir áreas a través del comportamiento alimentario, que con el tiempo despeja la vegetación, incluidas las plantas exóticas invasoras. La cantidad de semillas que esparcen los megaherbívoros es mayor que la de otros frugívoros. [4] [50] Además, los megaherbívoros que pastan, como el rinoceronte blanco, tienen un profundo impacto en la hierba corta. En un estudio, la hierba corta se volvió menos frecuente después de la eliminación de los rinocerontes blancos; este evento afectó a los herbívoros más pequeños que habían dependido de ellos para regular la población de hierba corta. [48]
En un estudio de 2018, se concluyó que los megaherbívoros no se veían afectados por el "paisaje del miedo", un paisaje en el que las presas evitan ciertas zonas de depredación críticas, alterando así las cascadas tróficas asustadas por los depredadores . Sus heces eran más visibles en áreas cerradas y densas, lo que indica que distribuyen recursos a áreas de riesgo en este "paisaje del miedo". [51]
Interacciones interespecíficas
La mayoría de las especies megaherbívoras son demasiado grandes y poderosas para que la mayoría de los depredadores las maten. [38] Sin embargo, las crías son el objetivo de varias especies de depredadores. [37] : 158 Las jirafas también son susceptibles a la depredación, y no es raro que los leones y las hienas manchadas cacen jirafas adultas en algunos lugares. Los jóvenes son especialmente vulnerables, ya que entre una cuarta parte y la mitad de las crías de jirafa no llegan a la edad adulta. [52] [53] En el Parque Nacional de Chobe , se ha registrado que los leones cazan elefantes jóvenes y subadultos. [54] Los tigres son otro depredador conocido de los elefantes jóvenes. [55] Las crías de hipopótamo a veces pueden ser presas de leones, hienas manchadas y cocodrilos del Nilo . [56]
Las jirafas pueden huir o actuar de manera no agresiva, mientras que los rinocerontes blancos no suelen reaccionar ante la presencia de depredadores. Los rinocerontes negros e indios, los elefantes y los hipopótamos, por otro lado, reaccionan con fuerza ante los depredadores. [37] : 124–131
Adaptaciones de los megaherbívoros actuales
Tamaño
Los elefantes son los miembros más grandes, con un peso de entre 2,5 y 6,0 toneladas. Los rinocerontes indios, los rinocerontes blancos y los hipopótamos suelen pesar entre 1,4 y 2,3 toneladas. El rinoceronte de Java y el rinoceronte negro tienen un peso medio de entre 1 y 1,3 toneladas. Las jirafas son los miembros más pequeños, con un rango de peso general de entre 0,8 y 1,2 toneladas. [37] : 14 [57]
Selección K
Los megaherbívoros actuales son especies seleccionadas por K , lo que significa que tienen altas expectativas de vida, crecimiento poblacional lento, crías grandes, embarazos prolongados y bajas tasas de mortalidad. Han seleccionado la reproducción lenta para mejorar sus posibilidades de supervivencia y, como resultado, aumentar su esperanza de vida. [58] [59] Su gran tamaño ofrece protección contra los depredadores, pero al mismo tiempo, disminuye el grado en el que se reproducen debido a las fuentes de alimento restringidas. [60] [61] Este lento crecimiento poblacional (los elefantes, por ejemplo, crecen a una tasa del 6-7 %), indica que las poblaciones pueden reducirse drásticamente si las presiones de depredación son demasiado grandes. [37] : 293 En entornos estables, la selección K predomina como la capacidad de competir con éxito por recursos limitados, y las poblaciones de organismos seleccionados por K suelen ser muy constantes en número y cercanas al máximo que el entorno puede soportar. [60] [37] : 200
Reproducción
_female_and_calf_(14007572496).jpg){kind=link}
_female_and_calf_(14007572496).jpg/1280px-Giraffes_(Giraffa_camelopardalis)_female_and_calf_(14007572496).jpg)
{kind=link}

Cuando las hembras entran en celo , los machos intentarán atraerlas y aparearse con ellas. Estas oportunidades de reproducción pueden estar influenciadas por el sistema jerárquico de los machos. Las jirafas y los elefantes se aparean durante un tiempo relativamente corto, mientras que los rinocerontes y los hipopótamos tienen una sesión de apareamiento que dura un período prolongado. Las hembras tienen largos períodos de gestación , entre 8 y 22 meses. Los intervalos entre los nacimientos varían entre las especies, pero el rango general es de 1,3 a 4,5 años. [37] : 116–124
Por lo general, dan a luz a una sola cría que depende en gran medida de las hembras para alimentarse y protegerse. A medida que crecen, la cría comienza a destetarse mientras aún está mamando. Cuando alcanzan la etapa juvenil, son capaces de valerse por sí mismas, pero solo hasta cierto punto. Las hembras suelen separarse de sus crías persiguiéndolas. A pesar de esto, las hembras pueden seguir interactuando con su progenie incluso después del destete. [37] : 133
Esperanza de vida y mortalidad
Los hipopótamos y los rinocerontes pueden vivir hasta 40 años, mientras que los elefantes pueden vivir más de 60 años. [53] Las jirafas tienen una vida útil de alrededor de 25 años. [37] : 158
Entre el 2 y el 5% de los megaherbívoros adultos mueren cada año. Los machos tienen más probabilidades que las hembras de morir por heridas sufridas durante las disputas. Ocasionalmente, en épocas de sequía, las poblaciones pueden reducirse significativamente, siendo las crías las más afectadas en esos períodos. [37] : 158
Véase también
- Megafauna australiana
- Gigantismo de las profundidades marinas
- Los animales más grandes y pesados
- Lista de megaherbívoros actuales
Referencias
- ^ ab "Megaherbívoro". Diccionario Collins . Consultado el 11 de enero de 2024.
Las praderas marinas tropicales sustentan una diversidad de herbívoros que abarcan las escalas meso, macro y megaherbívora.
- ^ ab Onstein, Renske E.; Kissling, W. Daniel; Linder, H. Peter (13 de abril de 2022). "La brecha de los megaherbívoros después de las extinciones de los dinosaurios no aviares modificó la evolución de los rasgos y la diversificación de las palmas tropicales". Actas de la Royal Society B: Ciencias Biológicas . 289 (1972). doi :10.1098/rspb.2021.2633. ISSN 0962-8452. PMC 9006001 . PMID 35414237.
- ^ abc Malhi, Yadvinder; Doughty, Christopher E.; Galetti, Mauro; Smith, Felisa A.; Svenning, Jens-Christian; Terborgh, John W. (25 de enero de 2016). "Megafauna y función del ecosistema desde el Pleistoceno hasta el Antropoceno". Actas de la Academia Nacional de Ciencias . 113 (4): 838–846. Bibcode :2016PNAS..113..838M. doi : 10.1073/pnas.1502540113 . ISSN 0027-8424. PMC 4743772 . PMID 26811442.
- ^ abcd Bocherens, Hervé (2018). "El ascenso de la antroposfera desde hace 50.000 años: ¿un reemplazo ecológico de megaherbívoros por humanos en ecosistemas terrestres?". Fronteras en ecología y evolución . 6 . doi : 10.3389/fevo.2018.00003 . ISSN 2296-701X.
- ^ ab Sulej, Tomasz; Niedźwiedzki, Grzegorz (4 de enero de 2019). "Un sinápsido del Triásico Tardío del tamaño de un elefante con extremidades erectas". Science . 363 (6422): 78–80. Bibcode :2019Sci...363...78S. doi :10.1126/science.aal4853. ISSN 0036-8075. PMID 30467179.
- ^ Viglietti, Pia A.; Benson, Roger BJ; Smith, Roger MH; Botha, Jennifer; Kammerer, Christian F.; Skosan, Zaituna; Butler, Elize; Crean, Annelise; Eloff, Bobby; Kaal, Sheena; Mohoi, Joël; Molehe, William; Mtalana, Nolusindiso; Mtungata, Sibusiso; Ntheri, Nthaopa (2021-04-27). "Evidencia de Sudáfrica de una extinción prolongada al final del Pérmico en tierra". Actas de la Academia Nacional de Ciencias . 118 (17). Bibcode :2021PNAS..11817045V. doi : 10.1073/pnas.2017045118 . ISSN 0027-8424. PMC 8092562 . Número de modelo: PMID33875588.
- ^ Fiorelli, Lucas E.; Ezcurra, Martín D.; Hechenleitner, E. Martín; Argañaraz, Eloísa; Taborda, Jeremías RA; Trotteyn, M. Jimena; von Baczko, M. Belén; Desojo, Julia B. (28 de noviembre de 2013). "Las letrinas comunitarias más antiguas conocidas proporcionan evidencia de gregarismo en los megaherbívoros del Triásico". Informes científicos . 3 (1): 3348. Código bibliográfico : 2013NatSR...3E3348F. doi :10.1038/srep03348. ISSN 2045-2322. PMC 3842779 . PMID 24287957.
- ^ Boitsova, Elizaveta A; Skutschas, Pavel P; Sennikov, Andrey G; Golubev, Valeriy K; Masuytin, Vladimir V; Masuytina, Olga A (5 de julio de 2019). "Histología ósea de dos pareiasaurios de Rusia (Deltavjatia rossica y Scutosaurus karpinskii) con implicaciones para la paleobiología de los pareiasaurios". Revista biológica de la Sociedad Linneana . doi :10.1093/biolinnean/blz094. ISSN 0024-4066.
- ^ Burgess, Seth D.; Bowring, Samuel A. (28 de agosto de 2015). "La geocronología de alta precisión confirma un magmatismo voluminoso antes, durante y después de la extinción más severa de la Tierra". Science Advances . 1 (7): e1500470. Bibcode :2015SciA....1E0470B. doi :10.1126/sciadv.1500470. ISSN 2375-2548. PMC 4643808 . PMID 26601239.
- ^ Hulse, D; Lau, KV; Sebastián, JV; Arndt, S; Meyer, KM; Ridgwell, A (28 de octubre de 2021). "Extinción marina del final del Pérmico debido al reciclaje de nutrientes impulsado por la temperatura y euxinia". Nat Geosci . 14 (11): 862–867. Código Bib : 2021NatGe..14..862H. doi :10.1038/s41561-021-00829-7. S2CID 240076553.
- ^ Wu, Yuyang; Chu, Daoliang; Tong, Jinnan; Song, Haijun; Dal Corso, Jacopo; Wignall, Paul B.; Song, Huyue; Du, Yong; Cui, Ying (9 de abril de 2021). "Aumento de seis veces del pCO2 atmosférico durante la extinción masiva del Pérmico-Triásico". Nature Communications . 12 (1): 2137. doi :10.1038/s41467-021-22298-7. ISSN 2041-1723. PMC 8035180 . PMID 33837195.
- ^ Erwin, Douglas H. (1990). "Patrones de diversidad de gasterópodos del Carbonífero-Triásico y la extinción masiva del Pérmico-Triásico". Paleobiología . 16 (2): 187–203. Bibcode :1990Pbio...16..187E. doi :10.1017/s0094837300009878. ISSN 0094-8373. S2CID 87789173.
- ^ Barras, Colin (8 de enero de 2024). «La extinción masiva que tal vez nunca hubiera ocurrido». New Scientist . Consultado el 8 de enero de 2024 .
- ^ Racki, Grzegorz; Lucas, Spencer G. (20 de abril de 2020). "Cronograma de extinción de los dicinodontes a la luz de una fauna polaca inusual del Triásico Tardío y el enfoque de Cuvier sobre la extinción". Biología histórica . 32 (4): 452–461. Bibcode :2020HBio...32..452R. doi :10.1080/08912963.2018.1499734. ISSN 0891-2963. S2CID 91926999.
- ^ Wyenberg-Henzler, Taia (2022). "Ocupación ecomorfoespacial de grandes dinosaurios herbívoros desde el Jurásico tardío hasta el Cretácico tardío en América del Norte". PeerJ . 10 : e13174. doi : 10.7717/peerj.13174 . ISSN 2167-8359. PMC 9009330 . PMID 35433123.
- ^ Button, David J; Barrett, Paul; Rayfield, Emily (noviembre de 2016). "Miología craneal comparativa y biomecánica de Plateosaurus y Camarasaurus y evolución del aparato de alimentación de los saurópodos". Paleontología . 59 (6): 887–913. Bibcode :2016Palgy..59..887B. doi :10.1111/pala.12266. ISSN 0031-0239. S2CID 53619160.
- ^ Grinham, Luke R. (agosto de 2023). "Aparición de megaherbívoros cenozoicos". Nature Ecology & Evolution . 7 (8): 1173. Bibcode :2023NatEE...7.1173G. doi :10.1038/s41559-023-02100-1. ISSN 2397-334X. PMID 37258656. S2CID 259000968.
- ^ Mallon, Jordan C.; Anderson, Jason S. (11 de junio de 2014). "Implicaciones funcionales y paleoecológicas de la morfología y el desgaste de los dientes de los dinosaurios megaherbívoros de la Formación Dinosaur Park (Campaniano superior) de Alberta, Canadá". PLOS ONE . 9 (6): e98605. Bibcode :2014PLoSO...998605M. doi : 10.1371/journal.pone.0098605 . ISSN 1932-6203. PMC 4053334 . PMID 24918431.
- ^ Brusatte, Stephen L.; Butler, Richard J.; Barrett, Paul M.; Carrano, Matthew T.; Evans, David C.; Lloyd, Graeme T.; Mannion, Philip D.; Norell, Mark A.; Peppe, Daniel J.; Upchurch, Paul; Williamson, Thomas E. (28 de julio de 2015). "La extinción de los dinosaurios". Biological Reviews . 90 (2): 628–642. doi :10.1111/brv.12128. hdl : 20.500.11820/176e5907-26ec-4959-867f-0f2e52335f88 . ISSN 1464-7931. PMID 25065505.
- ^ Clauss, M.; Frey, R.; Kiefer, B.; Lechner-Doll, M.; Loehlein, W.; Polster, C.; Rossner, GE; Streich, WJ (1 de junio de 2003). "El tamaño corporal máximo alcanzable de los mamíferos herbívoros: restricciones morfofisiológicas en el intestino anterior y adaptaciones de los fermentadores del intestino posterior". Oecologia . 136 (1): 14–27. Bibcode :2003Oecol.136...14C. doi :10.1007/s00442-003-1254-z. ISSN 0029-8549. PMID 12712314. S2CID 206989975.
- ^ Sanisidro, Oscar; Mihlbachler, Matthew C.; Cantalapiedra, Juan L. (12 de mayo de 2023). "Una vía macroevolutiva hacia la megaherbivoría". Science . 380 (6645): 616–618. Bibcode :2023Sci...380..616S. doi :10.1126/science.ade1833. ISSN 0036-8075. PMID 37167399. S2CID 258618428.
- ^ Domning, DP (febrero de 2001). "Sirenios, pastos marinos y cambio ecológico cenozoico en el Caribe". Paleogeografía, Paleoclimatología, Paleoecología . 166 (1–2): 27–50. Bibcode :2001PPP...166...27D. doi :10.1016/S0031-0182(00)00200-5.
- ^ Price, Gilbert J.; Fitzsimmons, Kathryn E.; Nguyen, Ai Duc; Zhao, Jian-xin; Feng, Yue-xing; Sobbe, Ian H.; Godthelp, Henk; Archer, Michael; Hand, Suzanne J. (30 de noviembre de 2021). "Nuevas edades del marsupial más grande del mundo: Diprotodon optatum de la Australia del Pleistoceno". Quaternary International . Evolución humana en el ámbito de Asia y el Pacífico: Actas de la 1.ª Conferencia de Asia y el Pacífico sobre la evolución humana. 603 : 64–73. Bibcode :2021QuInt.603...64P. doi :10.1016/j.quaint.2021.06.013. ISSN 1040-6182.
- ^ Palmer, D., ed. (1999). La enciclopedia ilustrada Marshall de dinosaurios y animales prehistóricos . Londres: Marshall Editions. pág. 208. ISBN 1-84028-152-9.
- ^ Saarinen, Juha; Karme, Aleksis (junio de 2017). "Desgaste dental y dietas de xenartros actuales y fósiles (Mammalia, Xenarthra): aplicación de un nuevo enfoque de desgaste mesofítico". Paleogeografía, Paleoclimatología, Paleoecología . 476 : 42–54. Bibcode :2017PPP...476...42S. doi :10.1016/j.palaeo.2017.03.027.
- ^ van Geel, Bas; van Leeuwen, Jacqueline FN; Nooren, Kees; Mol, Dick; den Ouden, Natasja; van der Knaap, Pim WO; Seersholm, Frederik V.; Rey-Iglesia, Alba; Lorenzen, Eline D. (enero de 2022). "Dieta y ambiente de Mylodon darwinii basada en polen de un coprolito del Glaciar Tardío de la Cueva del Mylodon en el sur de Chile". Revista de Paleobotánica y Palinología . 296 : 104549. Código Bib : 2022RPaPa.29604549V. doi : 10.1016/j.revpalbo.2021.104549 . S2CID 239902623.
- ^ Pujos, François; Gaudin, Timothy J.; De Iuliis, Gerardo; Cartelle, Cástor (septiembre de 2012). "Avances recientes en variabilidad, adaptaciones morfofuncionales, terminología dental y evolución de los perezosos". Journal of Mammalian Evolution . 19 (3): 159–169. doi :10.1007/s10914-012-9189-y. hdl : 11336/69736 . ISSN 1064-7554. S2CID 254701351.
- ^ Pérez-Crespo, VCAN; Arroyo-Cabrales, JN; Benammi, M.; Johnson, E.; Polaco, DO; Santos-Moreno, A.; Morales-Puente, P.; Cienfuegos-Alvarado, E. (2012). "Variación geográfica de la dieta y hábitat de las poblaciones mexicanas de mamut colombiano ( Mammuthus columbi )". Cuaternario Internacional . 276–277: 8–16. Código Bib : 2012QuiInt.276....8P. doi : 10.1016/j.quaint.2012.03.014.
- ^ Koch, PL; Barnosky, AD (2006). "Extinciones del Cuaternario tardío: estado del debate". Revista anual de ecología, evolución y sistemática . 37 : 215–250. doi :10.1146/annurev.ecolsys.34.011802.132415.
- ^ Gill, Jacquelyn L. (2014). "Impactos ecológicos de las extinciones de megaherbívoros del Cuaternario tardío". New Phytologist . 201 (4): 1163–1169. doi :10.1111/nph.12576. ISSN 0028-646X. PMID 24649488.
- ^ Martin, PS (1963). Los últimos 10.000 años: un registro de polen fósil del suroeste americano . Tucson, AZ: Univ. Ariz. Press. págs. 61–86. ISBN 978-0-8165-1759-6.
- ^ Martin, PS (1967). "Exceso de destrucción prehistórica". En Martin, PS; Wright, HE (eds.). Extinciones del Pleistoceno: la búsqueda de una causa . New Haven: Yale Univ. Press. págs. 75–120. ISBN 978-0-300-00755-8.
- ^ Martin, PS (1989). "Extinción prehistórica: un modelo global". En Martin, PS; Klein, RG (eds.). Extinciones cuaternarias: una revolución prehistórica . Tucson, AZ: University of Arizona Press. págs. 354–404. ISBN 978-0-8165-1100-6.
- ^ Birks, HH (1973). "Conjuntos de macrofósiles modernos en sedimentos lacustres de Minnesota". En Birks, HJB; West, RG (eds.). Ecología vegetal cuaternaria: el 14.º simposio de la British Ecological Society, Universidad de Cambridge, 28-30 de marzo de 1972. Oxford: Blackwell Scientific. ISBN 0-632-09120-7.
- ^ ab 'Wildlife' Review. Departamento del Interior de los Estados Unidos, Servicio de Pesca y Vida Silvestre. 1989.
- ^ Wells, Harry BM; Crego, Ramiro D.; Opedal, Øystein H.; Khasoha, Leo M.; Alston, Jesse M.; Reed, Courtney G.; Weiner, Sarah; Kurukura, Samson; Hassan, Abdikadir A.; Namoni, Mathew; Ekadeli, Jackson; Kimuyu, Duncan M.; Young, Truman P.; Kartzinel, Tyler R.; Palmer, Todd M. (2021). "Evidencia experimental de que los efectos de los megaherbívoros en el uso del espacio por parte de los mesoherbívoros están influenciados por los rasgos de las especies". Revista de Ecología Animal . 90 (11): 2510–2522. Código Bibliográfico :2021JAnEc..90.2510W. doi : 10.1111/1365-2656.13565 . Código IATA : 10 ...
- ^ abcdefghijklmn Owen-Smith, R. Norman (1988). Megaherbívoros: la influencia del tamaño corporal muy grande en la ecología. Cambridge University Press. ISBN 978-0-521-42637-4.
- ^ abc Ray, Justina; Redford, Kent H.; Steneck, Robert; Berger, Joel (9 de abril de 2013). Grandes carnívoros y la conservación de la biodiversidad. Island Press. pág. 87. ISBN 978-1-59726-609-3.
- ^ Gheerbrant, E. (2009). "Aparición de parientes de los elefantes en el Paleoceno y la rápida radiación de ungulados africanos". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América . 106 (26): 10717–10721. Bibcode :2009PNAS..10610717G. doi : 10.1073/pnas.0900251106 . PMC 2705600 . PMID 19549873.
- ^ Gatesy, J. (1 de mayo de 1997). "Más apoyo de ADN para un clado Cetacea/Hippopotamidae: el gen de la proteína de coagulación sanguínea gamma-fibrinógeno". Biología molecular y evolución . 14 (5): 537–543. doi : 10.1093/oxfordjournals.molbev.a025790 . PMID 9159931.
- ^ Boisserie, Jean-Renaud; Lihoreau, Fabrice; Brunet, Michel (2005). "La posición de Hippopotamidae dentro de Cetartiodactyla". Actas de la Academia Nacional de Ciencias . 102 (5): 1537–1541. Bibcode :2005PNAS..102.1537B. doi : 10.1073/pnas.0409518102 . PMC 547867 . PMID 15677331.
- ^ Chen, Lei; Qiu, Qiang; Jiang, Yu; Wang, Kun; Lin, Zeshan; et al. (2019-06-21). "La secuenciación del genoma de los rumiantes a gran escala proporciona información sobre su evolución y sus rasgos distintivos". Science . 364 (6446). Bibcode :2019Sci...364.6202C. doi :10.1126/science.aav6202. ISSN 0036-8075. PMID 31221828.
- ^ ab Ripple, William J.; Newsome, Thomas M.; Wolf, Christopher; Dirzo, Rodolfo; Everatt, Kristoffer T.; et al. (mayo de 2015). "Colapso de los herbívoros más grandes del mundo". Science Advances . 1 (4): e1400103. Bibcode :2015SciA....1E0103R. doi :10.1126/sciadv.1400103. ISSN 2375-2548. PMC 4640652 . PMID 26601172.
- ^ Schowanek, Simón D.; Davis, Matt; Lundgren, Erick J.; Middleton, Owen; Serbal, Juan; Pedersen, Rasmus Ø.; Rampa, Daniel; Sandom, Christopher J.; Svenning, Jens-Christian (abril de 2021). Lyon, Kathleen (ed.). "La reintroducción de herbívoros extirpados podría revertir parcialmente el declive del Cuaternario tardío de especies grandes y de pastoreo". Ecología Global y Biogeografía . 30 (4): 896–908. Código Bib : 2021GloEB..30..896S. doi :10.1111/geb.13264. ISSN 1466-822X. S2CID 233926684.
- ^ Owen-Smith, N. (2013). "Megaherbívoros". Enciclopedia de la biodiversidad (segunda edición). págs. 223-229. doi :10.1016/B978-0-12-384719-5.00358-0. ISBN 9780123847201.
- ^ Clauss, Marcus; Frey, R; Kiefer, B; Lechner-Doll, M. (2003). "El tamaño corporal máximo alcanzable de los mamíferos herbívoros: restricciones morfofisiológicas en el intestino anterior y adaptaciones de los fermentadores del intestino posterior". Oecologia . 136 (1): 14–27. Bibcode :2003Oecol.136...14C. doi :10.1007/s00442-003-1254-z. PMID 12712314. S2CID 206989975.
- ^ Ruggiero, Richard G. (marzo de 1991). "Depredación oportunista de crías de elefante". Revista Africana de Ecología . 29 (1): 86–89. Código Bibliográfico :1991AfJEc..29...86R. doi :10.1111/j.1365-2028.1991.tb00823.x. ISSN 0141-6707.
- ^ ab Hyvärinen, Olli (2022). Megaherbívoros y funcionamiento del sistema terrestre: efectos a escala del paisaje de la pérdida del rinoceronte blanco en la vegetación, los incendios y la dinámica del carbono del suelo. Universidad Sueca de Ciencias Agrícolas. p. 107. ISBN 978-91-7760-966-7.
- ^ Gill, Jacquelyn L. (2013). "Impactos ecológicos de las extinciones de megaherbívoros del Cuaternario tardío". New Phytologist . 201 (4): 1163–1169. doi :10.1111/nph.12576. PMID 24649488.
- ^ Mungi, Ninad Avinash; Jhala, Yadvendradev V.; Qureshi, Qamar; le Roux, Elizabeth; Svenning, Jens-Christian (marzo de 2023). "Los megaherbívoros proporcionan resistencia biótica contra el dominio de plantas exóticas". Ecología y evolución de la naturaleza . 7 (10): 1645-1653. doi :10.1038/s41559-023-02181-y. ISSN 2397-334X.
- ^ le Roux, Elizabeth; Kerley, Graham IH; Cromsigt, Joris PGM (agosto de 2018). "Los megaherbívoros modifican las cascadas tróficas desencadenadas por el miedo a la depredación en un ecosistema de sabana africana". Current Biology . 28 (15): 2493–2499.e3. doi : 10.1016/j.cub.2018.05.088 . ISSN 0960-9822. PMID 30033334.
- ^ Lee, Derek E.; Bond, Monica L.; Kissui, Bernard M.; Kiwango, Yustina A.; Bolger, Douglas T. (11 de mayo de 2016). "Variación espacial en la demografía de las jirafas: una prueba de dos paradigmas". Revista de mastozoología . 97 (4): 1015–1025. doi :10.1093/jmammal/gyw086. ISSN 1545-1542.
- ^ ab Bruton, Michael N. (6 de diciembre de 2012). Estilos alternativos de historia de vida de los animales. Springer Science & Business Media. pág. 447. ISBN 978-94-009-2605-9.
- ^ Power, R. John; Shem Compion, RX (abril de 2009). "Depredación de elefantes por parte de leones en Savuti, Parque Nacional Chobe, Botsuana". Zoología africana . 44 (1): 36–44. doi :10.3377/004.044.0104. ISSN 1562-7020. S2CID 86371484.
- ^ Thuppil, Vivek; Coss, Richard G. (23 de octubre de 2013). "Los elefantes asiáticos salvajes distinguen los gruñidos agresivos de tigres y leopardos según el peligro percibido". Biology Letters . 9 (5): 20130518. doi :10.1098/rsbl.2013.0518. ISSN 1744-9561. PMC 3971691 . PMID 24026347.
- ^ Estes, R. (1992). Guía de comportamiento de los mamíferos africanos: incluye mamíferos ungulados, carnívoros y primates. University of California Press. pp. 222–226. ISBN 978-0-520-08085-0.
- ^ Larramendi, A. (2016). "Altura de los hombros, masa corporal y forma de los proboscídeos" (PDF) . Acta Palaeontologica Polonica . 61 (3): 537–574. doi :10.4202/app.00136.2014. ISSN 0888-8892. S2CID 84294169.
- ^ Manlik, Oliver (2019). "La importancia de la reproducción para la conservación de poblaciones animales de crecimiento lento". Ciencias reproductivas en conservación animal . Avances en medicina y biología experimental. Vol. 1200. págs. 13–39. doi :10.1007/978-3-030-23633-5_2. ISBN . 978-3-030-23632-8. ISSN 0065-2598. PMID 31471793. S2CID 201756810.
- ^ Yuan, Rong; Hascup, Erin; Hascup, Kevin; Bartke, Andrzej (1 de noviembre de 2023). "Relaciones entre desarrollo, crecimiento, tamaño corporal, reproducción, envejecimiento y longevidad: compensaciones y ritmo de vida". Biochemistry (Moscú) . 88 (11): 1692–1703. doi : 10.1134/S0006297923110020 . ISSN: 1608-3040. PMC: 10792675. PMID : 38105191. S2CID : 265507842.
- ^ ab Rafferty, John P. "Especies seleccionadas por K". Britannica . Consultado el 2 de abril de 2017 .
- ^ Trimble, MJ; Ferreira, SM; Van Aarde, RJ (septiembre de 2009). "Impulsores de las fluctuaciones demográficas de los megaherbívoros: inferencia a partir de los elefantes". Revista de Zoología . 279 (1): 18–26. doi :10.1111/j.1469-7998.2009.00560.x. ISSN 0952-8369.
