Terápsidos
| Terápsidos Rango temporal: | |
|---|---|
 | |
| De arriba a abajo y de izquierda a derecha, varios ejemplos de terápsidos no mamíferos: Biarmosuchus ( Biarmosuchia ), Moschops ( Dinocephalia ), Lystrosaurus ( Anomodontia ), Inostrancevia ( Gorgonopsia ), Glanosuchus ( Therocephalia ) y Chiniquodon ( Cynodontia ). | |
| Clasificación científica | |
| Dominio: | Eucariota |
| Reino: | Animalia |
| Filo: | Cordados |
| Clado : | Sinápsidos |
| Clado : | Esfenacodoncia |
| Clado : | Pantera ápsida |
| Clado : | Esfenacodontoides |
| Clado : | Retama Therapsida , 1905 [1] |
| Clados | |
| |
{kind=link}
Therapsida [a] es un clado que comprende un grupo importante de sinápsidos eupelicosaurios que incluye a los mamíferos y sus ancestros y parientes cercanos. Muchos de los rasgos que hoy se consideran exclusivos de los mamíferos tuvieron su origen en los primeros terápsidos, incluidas las extremidades que estaban orientadas más hacia abajo del cuerpo, lo que resultó en una postura cuadrúpeda más "de pie" , en oposición a la postura agachada de muchos reptiles y anfibios .
Los terápsidos evolucionaron a partir de sinápsidos anteriores comúnmente llamados " pelicosaurios ", específicamente dentro de Sphenacodontia , hace más de 279,5 millones de años. Reemplazaron a los pelicosaurios como los animales terrestres grandes dominantes desde el Guadalupiano hasta el Triásico Inferior. Después de la extinción masiva del Pérmico-Triásico , los terápsidos perdieron importancia relativa frente a los saurópsidos arcosaurios ( pseudosuquios , dinosaurios y pterosaurios , etc.) que se diversificaron rápidamente durante el Triásico Medio.
Los terápsidos incluyen a los cinodontos , el grupo que dio origen a los mamíferos ( Mammaliaformes ) en el Triásico Tardío hace unos 225 millones de años, el único clado de terápsidos que sobrevivió más allá del final del Triásico . El único otro grupo de terápsidos que sobrevivió hasta el Triásico Tardío , los dicinodontos , se extinguieron hacia el final del período. El último grupo sobreviviente de cinodontos no mamaliaformes fueron los Tritylodontidae , que se extinguieron durante el Cretácico Inferior .
Características
{kind=link}

Mandíbula y dientes
Las fenestras temporales de los terápsidos eran más grandes que las de los pelicosaurios. Las mandíbulas de algunos terápsidos eran más complejas y poderosas, y los dientes se diferenciaban en incisivos frontales para pellizcar, grandes caninos laterales para perforar y desgarrar, y molares para cortar y picar los alimentos.
Postura
Las patas de los terápsidos estaban situadas más verticalmente debajo de sus cuerpos que las patas extendidas de los reptiles y pelicosaurios. Además, en comparación con estos grupos, los pies eran más simétricos, con el primer y el último dedo cortos y los dedos del medio largos, una indicación de que el eje del pie estaba colocado paralelo al del animal, no extendido hacia los lados. Esta orientación habría dado un andar más parecido al de los mamíferos que al de los lagartos de los pelicosaurios. [2]
Fisiología
La fisiología de los terápsidos es poco conocida. La mayoría de los terápsidos del Pérmico tenían un agujero pineal, lo que indica que tenían un ojo parietal como muchos reptiles y anfibios modernos. El ojo parietal cumple una función importante en la termorregulación y el ritmo circadiano de los ectotérmicos, pero está ausente en los mamíferos modernos, que son endotérmicos . [3] Cerca del final del Pérmico, los dicinodontos, los terocéfalos y los cinodontes muestran tendencias paralelas hacia la pérdida del agujero pineal, y el agujero está completamente ausente en los cinodontes probainognatios. La evidencia de los isótopos de oxígeno, que se correlacionan con la temperatura corporal, sugiere que la mayoría de los terápsidos del Pérmico eran ectotérmicos y que la endotermia evolucionó de manera convergente en los dicinodontos y cinodontes cerca del final del Pérmico. [4] Por el contrario, la evidencia de la histología sugiere que la endotermia es compartida por todos los Therapsida, [5] mientras que las estimaciones de la tasa de flujo sanguíneo y la esperanza de vida en el mamífero Morganucodon sugieren que incluso los primeros mamíferos tenían tasas metabólicas similares a las de los reptiles. [6] Se informó evidencia de cornetes respiratorios, que se ha planteado como indicativos de endotermia, en el terocéfalo Glanosuchus , pero un estudio posterior mostró que los aparentes sitios de unión de los cornetes pueden ser simplemente el resultado de la distorsión del cráneo. [7]
Integumento
La evolución del tegumento en los terápsidos es poco conocida, y hay pocos fósiles que proporcionen evidencia directa de la presencia o ausencia de pelo. Los sinápsidos más basales con evidencia directa inequívoca de pelo son los docodontes , que son mamíferos muy relacionados con los mamíferos del grupo corona. Dos especímenes juveniles "momificados" del dicinodonte Lystrosaurus murrayi conservan impresiones de piel; la piel no tiene pelo, es correosa y tiene hoyuelos, algo comparable a la piel de elefante. [8] [9] Se ha descrito que la piel facial fosilizada del dinocéfalo Estemmenosuchus muestra que la piel era glandular y carecía tanto de escamas como de pelo. [10]
Se han encontrado coprolitos que contienen lo que parecen ser pelos del Pérmico tardío . [11] [12] Aunque no se conoce con certeza el origen de estos pelos, pueden sugerir que el cabello estaba presente en al menos algunos terápsidos del Pérmico.
El cierre del agujero pineal en los cinodontos probainognatios puede indicar una mutación en el gen regulador Msx2, que está involucrado tanto en el cierre del techo del cráneo como en el mantenimiento de los folículos pilosos en ratones. [13] Esto sugiere que el cabello puede haber evolucionado primero en los probainognatios, aunque no descarta por completo un origen anterior del pelaje. [13]
Los bigotes probablemente evolucionaron en los cinodontos probainognatios. [13] [14] Algunos estudios habían inferido un origen anterior para los bigotes basándose en la presencia de agujeros en el hocico de los terocéfalos y los primeros cinodontos, pero la disposición de los agujeros en estos taxones en realidad se parece mucho a la de los lagartos, [15] lo que haría poco probable la presencia de bigotes parecidos a los de los mamíferos. [14]
Historia evolutiva
{kind=link}

Los terápsidos evolucionaron a partir de un grupo de pelicosaurios llamados esfenacodontos . [17] [18] Los terápsidos se convirtieron en los animales terrestres dominantes en el Pérmico medio , desplazando a los pelicosaurios. Los terápsidos constan de cuatro clados principales : los dinocéfalos , los anomodontes herbívoros , los biarmosuquios carnívoros y los teriodontes mayoritariamente carnívoros . Después de un breve estallido de diversidad evolutiva, los dinocéfalos se extinguieron a finales del Pérmico medio ( Guadalupiense ), pero los dicinodontes anomodontes , así como los gorgonopsianos y terocéfalos teriodontes florecieron, uniéndose a ellos al final del Pérmico el primero de los cinodontes .
{kind=link}

Al igual que todos los animales terrestres, los terápsidos se vieron gravemente afectados por el evento de extinción masiva del Pérmico-Triásico , con los muy exitosos gorgonopsios y los biarmosuquios muriendo por completo y los grupos restantes ( dicinodontos , terocéfalos y cinodontes) reducidos a un puñado de especies cada uno en el Triásico temprano . Los dicinodontos sobrevivientes estaban representados por dos familias de taxones de desastre ( Lystrosauridae y Myosauridae ), los escasamente conocidos Kombuisia y un solo grupo de herbívoros grandes y robustos , los Kannemeyeriiformes , que fueron el único linaje de dicinodontos que prosperó durante el Triásico. [20] Ellos y los cinodontes de tamaño mediano (incluyendo formas carnívoras y herbívoras) florecieron en todo el mundo durante el Triásico temprano y medio. Desaparecen del registro fósil en gran parte de Pangea al final del Carniano (Triásico tardío), aunque continuaron durante algún tiempo más en la franja ecuatorial húmeda y en el sur. [ cita requerida ]
Algunas excepciones fueron los eucinodontes , que derivaron aún más en la historia . [21] Al menos tres grupos de ellos sobrevivieron. Todos aparecieron en el período Triásico Tardío . La familia extremadamente parecida a los mamíferos , Tritylodontidae , sobrevivió hasta el Cretácico Inferior . Otra familia extremadamente parecida a los mamíferos, Tritheledontidae , es desconocida después del Jurásico Inferior . Mammaliaformes fue el tercer grupo, que incluía a Morganucodon y animales similares. Algunos taxónomos se refieren a estos animales como "mamíferos", aunque la mayoría limita el término al grupo de la corona de los mamíferos .
{kind=link}

Los cinodontos no eucinodontes sobrevivieron a la extinción del Pérmico-Triásico; Thrinaxodon , Galesaurus y Platycraniellus son conocidos del Triásico Inferior . Sin embargo, hacia el Triásico Medio , solo sobrevivieron los eucinodontes.
Los terocéfalos , parientes de los cinodontes, lograron sobrevivir a la extinción del Pérmico-Triásico y continuaron diversificándose durante el período Triásico Inferior. Sin embargo, al acercarse el final del período, los terocéfalos estaban en declive hasta su eventual extinción, probablemente superados por el linaje de los diápsidos , que se diversificaba rápidamente y estaba equipado con sistemas respiratorios sofisticados más adecuados para el mundo muy caluroso, seco y pobre en oxígeno del Triásico Final.
Los dicinodontos estuvieron entre los grupos de terápsidos más exitosos durante el Pérmico tardío y sobrevivieron hasta casi el final del Triásico.
Los mamíferos son los únicos terápsidos vivos. El grupo de la corona de los mamíferos , que evolucionó a principios del período Jurásico , se originó a partir de un grupo de mamíferos formadores que incluía a los docodontes . Los mamíferos formadores a su vez evolucionaron a partir de los probainognatios , un linaje del suborden eucinodonte.
Clasificación
| El paradigma de Hopson y Barghausen para las relaciones terápsidas |
Se reconocen generalmente seis grupos principales de terápsidos: Biarmosuchia , Dinocephalia , Anomodontia , Gorgonopsia , Therocephalia y Cynodontia . Un clado que une a los terocéfalos y los cinodontes, llamado Eutheriodontia , está bien respaldado, pero las relaciones entre los otros cuatro clados son controvertidas. [23] La hipótesis más ampliamente aceptada de las relaciones de los terápsidos, el paradigma de Hopson y Barghausen, fue propuesta por primera vez en 1986. Según esta hipótesis, los biarmosuquios son el grupo principal de terápsidos que diverge más temprano, y los otros cinco grupos forman Eutherapsida, y dentro de Eutherapsida, los gorgonopsianos son el taxón hermano de los eutheriodontos, que juntos forman Theriodontia . Hopson y Barghausen no llegaron inicialmente a una conclusión sobre cómo se relacionaban entre sí los dinocéfalos, los anomodontos y los teriodontes, pero estudios posteriores sugirieron que los anomodontos y los teriodontes deberían clasificarse juntos como Neotherapsida. Sin embargo, sigue habiendo debate sobre estas relaciones; en particular, algunos estudios han sugerido que los anomodontos, no los gorgonopsianos, son el taxón hermano de Eutheriodontia, otros estudios han descubierto que los dinocéfalos y los anomodontos forman un clado, y tanto la posición filogenética como la monofilia de Biarmosuchia siguen siendo controvertidas.
Además de los seis grupos principales, existen otros linajes y especies de clasificación incierta. Es probable que Raranimus, del Pérmico medio temprano de China, sea el terápsido conocido que divergió más temprano. [24] Se ha planteado la hipótesis de que Tetraceratops, del Pérmico temprano de los Estados Unidos, sea un terápsido que divergió aún más temprano, [25] [26] pero estudios más recientes han sugerido que es más probable que sea un esfenacodóntido no terápsido. [27]
Biarmosuquia
{kind=link}

Biarmosuchia es el clado de terápsidos reconocido más recientemente, reconocido por primera vez como un linaje distinto por Hopson y Barghausen en 1986 y nombrado formalmente por Sigogneau-Russell en 1989. La mayoría de los biarmosuquios fueron clasificados previamente como gorgonopsianos. Biarmosuchia incluye al distintivo Burnetiamorpha , pero el apoyo a la monofilia de Biarmosuchia es relativamente bajo. Muchos biarmosuquios son conocidos por su extensa ornamentación craneal.
Dinocefalia
{kind=link}

La dinocefalia comprende dos grupos distintivos, los Anteosauria y los Tapinocefalia .
Históricamente, los dinocéfalos carnívoros, incluidos tanto los anteosaurios como los titanosúquidos, se llamaban titanosúquios y se clasificaban como miembros de Theriodontia, mientras que los herbívoros Tapinocephalidae se clasificaban como miembros de Anomodontia.
Anomodoncia
{kind=link}
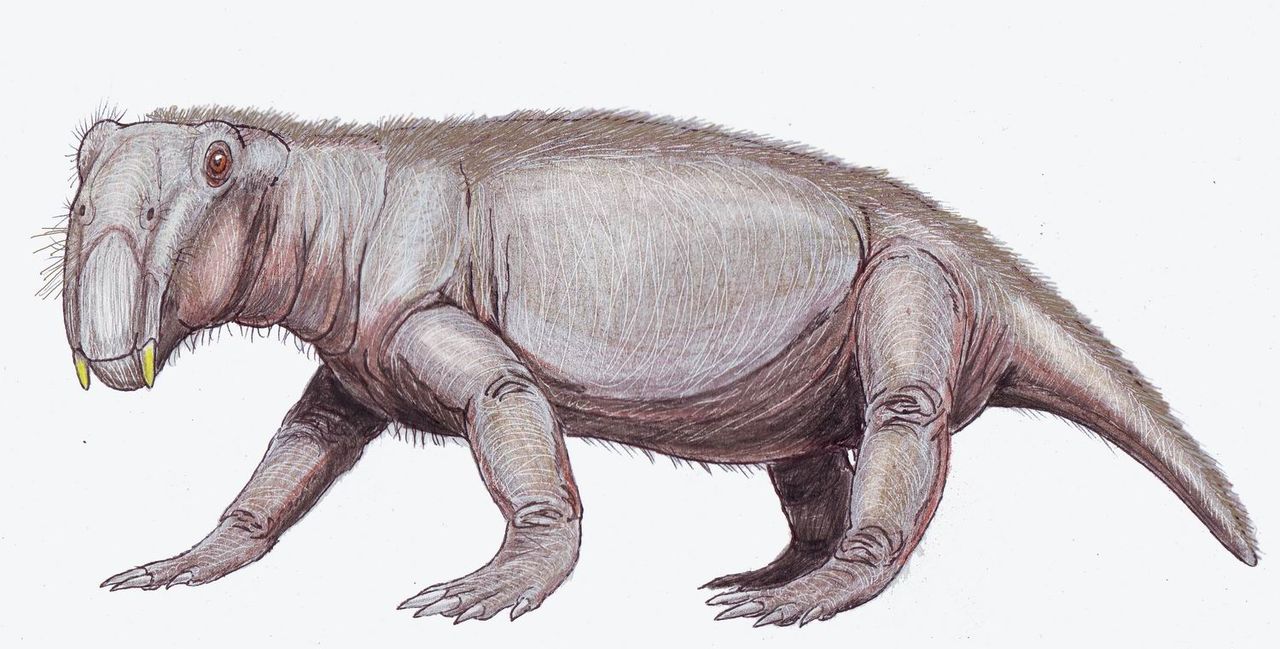
La anomodontia incluye a los dicinodontos , un clado de herbívoros con colmillos y pico, y el clado más diverso y longevo de terápsidos no cinodontes. Otros miembros de la anomodontia incluyen a Suminia , que se cree que era una forma trepadora.
Gorgonopsia
{kind=link}

Gorgonopsia es un grupo abundante pero morfológicamente homogéneo de depredadores dientes de sable .
Terócefalia
{kind=link}

Se ha sugerido que Therocephalia podría no ser monofilética, con algunas especies más estrechamente relacionadas con los cinodontos que otras. [28] Sin embargo, la mayoría de los estudios consideran a Therocephalia como monofilética.
Cinodontia
{kind=link}

Los cinodontos son el grupo de terápsidos más diverso y más longevo, ya que Cynodontia incluye a los mamíferos . Los cinodontos son el único clado importante de terápsidos que carece de un registro fósil del Pérmico medio, siendo el cinodonte más antiguo conocido Charassognathus de la era Wuchiapingiana del Pérmico tardío. Los cinodontos no mamíferos incluyen formas tanto carnívoras como herbívoras.
Véase también
- Evolución de los mamíferos
- Cronología de la historia evolutiva de la vida
- Paleontología de vertebrados
Notas
- ^ Griego : 'arco-bestia'
Referencias
- ^ Broom, R. (1905). "Sobre el uso del término Anomodontia". Registros del Museo de Albany . 1 (4): 266–269.
- ^ Carroll, RL (1988). Paleontología y evolución de vertebrados . Nueva York: WH Freeman and Company. pp. 698. ISBN. 978-0-7167-1822-2.
- ^ Benoit, Julien; Abdala, Fernando; Manger, Paul; Rubidge, Bruce (2016). "El sexto sentido en los precursores de los mamíferos: variabilidad del foramen parietal y la evolución del ojo pineal en los terápsidos euteriodontes del Pérmico-Triásico sudafricanos". Acta Palaeontologica Polonica . doi : 10.4202/app.00219.2015 . ISSN 0567-7920. S2CID 59143925.
- ^ Rey, Kevin; Amiot, Romain; Fourel, François; Abdalá, Fernando; Fluteau, Federico; Jalil, Nour-Eddine; Liu, junio; Rubidge, Bruce S; Smith, Roger MH; Steyer, J Sébastien; Viglietti, Pía A; Wang, Xu; Lécuyer, Christophe (18 de julio de 2017). "Los isótopos de oxígeno sugieren un termometabolismo elevado dentro de múltiples clados terápsidos del Permo-Triásico". eVida . 6 : –28589. doi : 10.7554/eLife.28589 . ISSN 2050-084X. PMC 5515572 . PMID 28716184.
- ^ Faure-Brac, Mathieu G.; Cubo, Jorge (2 de marzo de 2020). "¿Los sinápsidos eran primitivamente endotermos? Un enfoque paleohistológico utilizando mapas de vectores propios filogenéticos". Philosophical Transactions of the Royal Society B: Biological Sciences . 375 (1793): 20190138. doi :10.1098/rstb.2019.0138. ISSN 1471-2970. PMC 7017441 . PMID 31928185.
- ^ Newham, Elis; Gill, Pamela G.; Cervecero, Philippa; Benton, Michael J.; Fernández, Vicente; Gostling, Neil J.; Haberthür, David; Jernvall, Jukka; Kankaanpää, Tuomas; Kallonen, Aki; Navarro, Carlos; Pacureanu, Alexandra; Richards, Kelly; Marrón, Kate Robson; Schneider, Philipp; Suhonen, Heikki; Tafforeau, Paul; Williams, Katherine A.; Zeller-Plumhoff, Berit; Corfe, Ian J. (2020). "Fisiología similar a la de los reptiles en los mamíferos del tallo del Jurásico temprano". Comunicaciones de la naturaleza . 11 (1): 5121. Código bibliográfico : 2020NatCo..11.5121N. doi :10.1038/s41467-020-18898-4. ISSN 2041-1723 . PMC 7550344. PMID 33046697.
- ^ Hopson, James A. (18 de octubre de 2012). "El papel del modo de alimentación en el origen de los terápsidos: implicaciones para el origen de la endotermia de los mamíferos". Fieldiana Life and Earth Sciences . 5 : 126–148. doi :10.3158/2158-5520-5.1.126. ISSN 2158-5520. S2CID 84471370.
- ^ Smith, Roger MH; Botha, Jennifer; Viglietti, Pia A. (15 de octubre de 2022). "Tafonomía de los tetrápodos afectados por la sequía en la cuenca del Karoo del Triásico temprano, Sudáfrica". Paleogeografía, Paleoclimatología, Paleoecología . 604 : 111207. Bibcode :2022PPP...60411207S. doi :10.1016/j.palaeo.2022.111207.
- ^ Timmons, Jeanne (20 de septiembre de 2022). «Estas momias fósiles revelan un mundo brutal mucho antes de que viviera el T. Rex». Gizmodo . Consultado el 25 de julio de 2024 .
- ^ Chudinov, PK (1968). "Estructura de los tegumentos de los teromorfos". Doklady Akad. Nauk SSSR . 179 : 226–229.
- ^ Smith, Roger MH; Botha-Brink, Jennifer (2011). "Morfología y composición de coprolitos portadores de huesos del Grupo Beaufort del Pérmico Tardío, Cuenca Karoo, Sudáfrica". Paleogeografía, Paleoclimatología, Paleoecología . 312 (1–2): 40–53. Bibcode :2011PPP...312...40S. doi :10.1016/j.palaeo.2011.09.006. ISSN 0031-0182.
- ^ Bajdek, Piotr; Qvarnström, Martín; Owocki, Krzysztof; Sulej, Tomasz; Sennikov, Andrey G.; Golubev, Valeriy K.; Niedźwiedzki., Grzegorz (2016). "Microbiota y residuos de alimentos, incluida posible evidencia de pelo premamífero en coprolitos del Pérmico superior de Rusia". Lethaia . 49 (4): 455–477. Código Bib : 2016 Letha..49..455B. doi :10.1111/let.12156.
- ^ abc Benoit, J.; Manger, PR; Rubidge, BS (9 de mayo de 2016). "Pistas paleonurológicas sobre la evolución de los rasgos definitorios de los tejidos blandos de los mamíferos". Scientific Reports . 6 (1): 25604. Bibcode :2016NatSR...625604B. doi :10.1038/srep25604. ISSN 2045-2322. PMC 4860582 . PMID 27157809.
- ^ ab Benoit, Julien; Ruf, Irina; Miyamae, Juri A.; Fernandez, Vincent; Rodrigues, Pablo Gusmão; Rubidge, Bruce S. (2020). "La evolución del canal maxilar en Probainognathia (Cynodontia, Synapsida): reevaluación de la homología del foramen infraorbitario en ancestros mamíferos". Revista de evolución de mamíferos . 27 (3): 329–348. doi :10.1007/s10914-019-09467-8. ISSN 1573-7055. S2CID 156055693.
- ^ Estes, Richard (1961). "Anatomía craneal del reptil cinodonte Thrinaxodon liorhinus". Boletín del Museo de Zoología Comparada . 125 : 165–180.
- ^ Liu, Jun; Rubidge, Bruce; y Li, Jinling (2009). "Nuevo sinápsido basal apoya el origen laurasiático de los terápsidos" (PDF) . Acta Palaeontologica Polonica . 54 (3): 393–400. doi : 10.4202/app.2008.0071 . Consultado el 25 de septiembre de 2009 .
- ^ Clasificación de sinápsidos y apomorfías
- ^ Huttenlocker, Adam K. y Rega, Elizabeth (2012). "Capítulo 4. Paleobiología y microestructura ósea de los sinápsidos de grado pelicosaurio". En Chinsamy-Turan, Anusuya (ed.). Precursores de los mamíferos: radiación, histología, biología . Indiana University Press . págs. 90–119. ISBN. 978-0253005335.
- ^ Benoit, Julien; Norton, Luke A.; Manger, Paul R.; y Rubidge, Bruce S. (2017). Claessens, Leon (ed.). "Reevaluación de la capacidad de envenenamiento de Euchambersia mirabilis (Therapsida, Therocephalia) utilizando técnicas de escaneo μCT". PLOS ONE . 12 (2): e0172047. Bibcode :2017PLoSO..1272047B. doi : 10.1371/journal.pone.0172047 . PMC 5302418 . PMID 28187210.
- ^ Kammerer, Christian F.; Frobisch, Jörg; Angielczyk, Kenneth D. (31 de mayo de 2013). Farke, Andrew A. (ed.). "Sobre la validez y posición filogenética de Eubrachiosaurus browni, un dicinodonte Kannemeyeriiforme (Anomodontia) del Triásico de América del Norte". MÁS UNO . 8 (5): e64203. Código Bib : 2013PLoSO...864203K. doi : 10.1371/journal.pone.0064203 . ISSN 1932-6203. PMC 3669350 . PMID 23741307.
- ^ Padian, Kevin (4 de septiembre de 2013). "Una reseña de 'Precursores de los mamíferos: radiación, histología, biología'"". Revista de Paleontología de Vertebrados . 33 (5): 1250–1251. Código Bibliográfico :2013JVPal..33.1250P. doi :10.1080/02724634.2013.763814. ISSN 0272-4634.
- ^ Geggel, L. (2016). "Conoce al antiguo reptil que dio origen a los mamíferos". Scientific American .
- ^ Angielczyk, Kenneth D.; Kammerer, Christian F. (22 de octubre de 2018). "Sinápsidos no mamíferos: las raíces profundas del árbol genealógico de los mamíferos". En Zachos, Frank; Asher, Robert (eds.). Evolución, diversidad y sistemática de los mamíferos. De Gruyter. págs. 117–198. doi :10.1515/9783110341553-005. ISBN . 978-3-11-034155-3.S2CID 92370138 .
- ^ Duhamel, A.; Benoit, J.; Rubidge, BS y Liu, J. (agosto de 2021). "Una reevaluación del terápsido más antiguo, Raranimus, confirma su condición de miembro basal del clado y llena el vacío de Olson". La ciencia de la naturaleza . 108 (4): 26. Bibcode :2021SciNa.108...26D. doi :10.1007/s00114-021-01736-y. ISSN 0028-1042. PMID 34115204. S2CID 235403632.
- ^ Laurin, M.; Reisz, RR (1996). "La osteología y las relaciones de Tetraceratops insignis , el terápsido más antiguo conocido". Revista de Paleontología de Vertebrados . 16 (1): 95–102. Bibcode :1996JVPal..16...95L. doi :10.1080/02724634.1996.10011287.
- ^ Amsón, Elí; Laurín, Michel (2011). "Sobre las afinidades de Tetraceratops insignis, un sinápsido del Pérmico temprano". Acta Paleontológica Polonica . 56 (2): 301–312. doi : 10.4202/aplicación.2010.0063 . ISSN 0567-7920. S2CID 56425905.
- ^ Spindler, Frederik (2020). "El cráneo de Tetraceratops insignis (Synapsida, Sphenacodontia)". Paleovertebrados . 43 (1): e1. doi :10.18563/pv.43.1.e1. S2CID 214247325.
- ^ Abdalá, Fernando; Rubidge, Bruce S.; van den Heever, Juri (2008). "Los terocéfalos más antiguos (Therapsida, Eutheriodontia) y la diversificación temprana de Therapsida". Paleontología . 51 (4): 1011–1024. Código Bib : 2008Palgy..51.1011A. doi : 10.1111/j.1475-4983.2008.00784.x . ISSN 1475-4983. S2CID 129791548.
Lectura adicional
- Benton, MJ (2004). Paleontología de vertebrados , 3.ª ed., Blackwell Science.
- Carroll, RL (1988). Paleontología y evolución de vertebrados . WH Freeman & Company, Nueva York.
- Kemp, TS (2005). El origen y la evolución de los mamíferos . Oxford University Press .
- Romer, AS (1966). Paleontología de vertebrados . University of Chicago Press, 1933; 3.ª ed.
- Bennett, AF y Ruben, JA (1986). "El estado metabólico y termorregulador de los terápsidos". En Ecología y biología de reptiles parecidos a mamíferos . Smithsonian Institution Press, Washington, DC, 207-218.
- Kammerer, Christian F.; Angielczyk, Kenneth D.; Fröbisch, Jörg (2014). Historia evolutiva temprana de los sinápsidos. Paleobiología y paleoantropología de vertebrados. Bibcode :2014eehs.book.....K. doi :10.1007/978-94-007-6841-3. ISBN 978-94-007-6840-6.
- Ross, RP, Ross, CA (2023). Período Pérmico, geocronología . Enciclopedia Británica. https://www.britannica.com/science/Permian-Period
- Padian, Kevin (2013-09). "Una revisión de "Precursores de los mamíferos: radiación, histología, biología"". Revista de paleontología de vertebrados . 33 (5): 1250–1251.
Enlaces externos
- "Therapsida: Mamíferos y parientes extintos" El árbol de la vida
- "Therapsida: visión general" Paleos
