El impacto humano en el ciclo del nitrógeno
 | Este artículo tiene varios problemas. Ayúdenos a mejorarlo o a discutir estos problemas en la página de discusión . ( Aprenda cómo y cuándo eliminar estos mensajes )
|

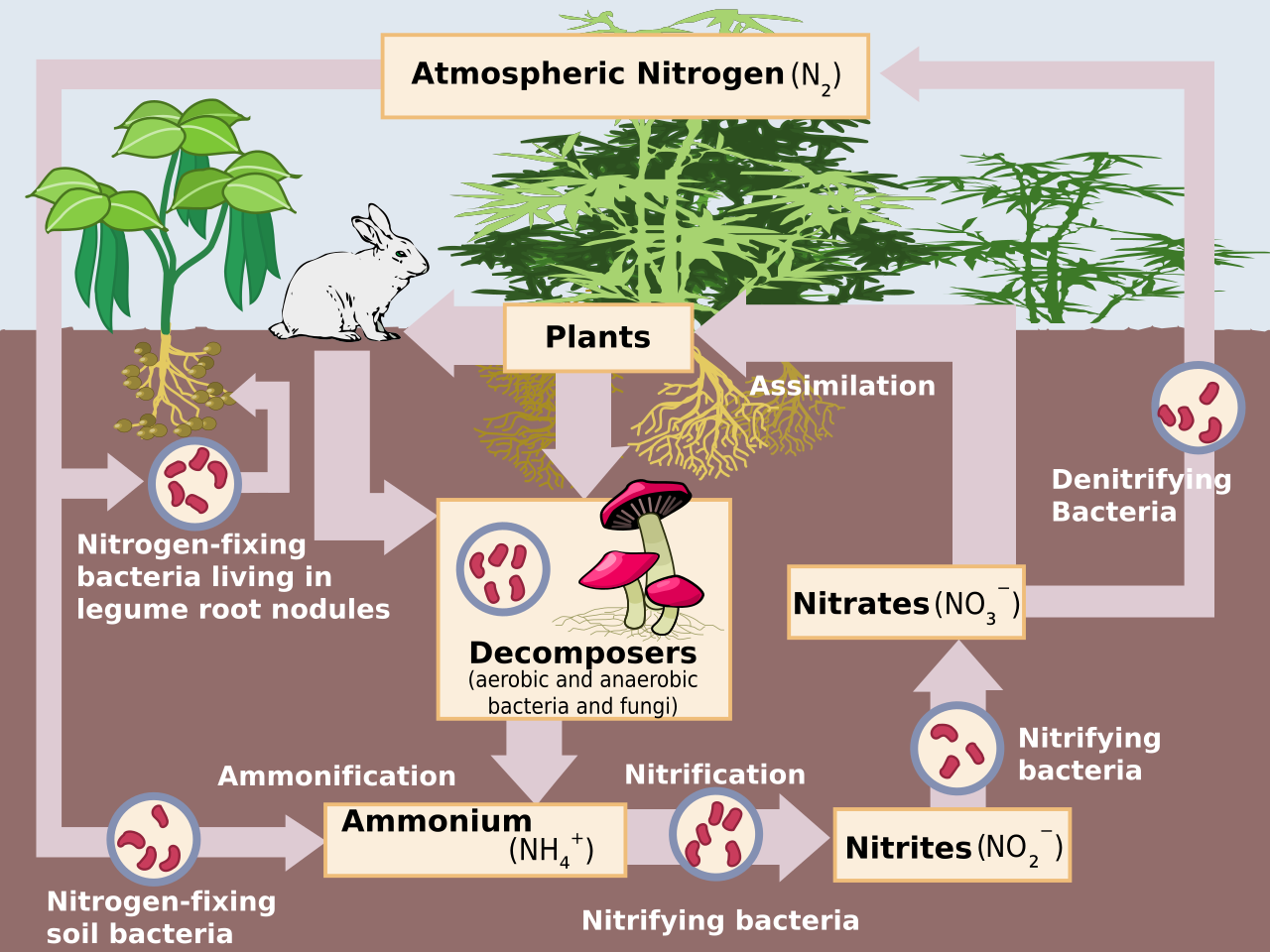

El impacto humano en el ciclo del nitrógeno es diverso. Los aportes de nitrógeno (N) agrícolas e industriales al medio ambiente actualmente superan los aportes de la fijación natural de N. [1] Como consecuencia de los aportes antropogénicos, el ciclo global del nitrógeno (Fig. 1) se ha alterado significativamente durante el siglo pasado. Las fracciones molares de óxido nitroso (N 2 O) atmosférico global han aumentado de un valor preindustrial de ~270 nmol/mol a ~319 nmol/mol en 2005. [2] Las actividades humanas representan más de un tercio de las emisiones de N 2 O, la mayoría de las cuales se deben al sector agrícola. [2] Este artículo tiene como objetivo brindar una breve revisión de la historia de los aportes antropogénicos de N y los impactos informados de los aportes de nitrógeno en ecosistemas terrestres y acuáticos seleccionados .
Historia de los aportes de nitrógeno antropogénico
| Tipos de alimentos | Emisiones acidificantes (g SO 2 eq por 100g de proteína) |
|---|---|
| Carne de res | |
| Queso | |
| Cerdo | |
| Cordero y carnero | |
| Crustáceos de cultivo | |
| Aves de corral | |
| Peces de cultivo | |
| Huevos | |
| Cacahuetes | |
| Guisantes | |
| Tofu |
Aproximadamente el 78% de la atmósfera de la Tierra es gas N (N 2 ), que es un compuesto inerte y biológicamente no disponible para la mayoría de los organismos. Para ser utilizado en la mayoría de los procesos biológicos, el N 2 debe convertirse en nitrógeno reactivo (Nr), que incluye formas inorgánicas reducidas (NH 3 y NH 4 + ), formas inorgánicas oxidadas (NO, NO 2 , HNO 3 , N 2 O y NO 3 − ) y compuestos orgánicos ( urea , aminas y proteínas ). [1] El N 2 tiene un triple enlace fuerte, por lo que se requiere una cantidad significativa de energía (226 kcal mol −1 ) para convertir el N 2 en Nr. [1] Antes de los procesos industriales, las únicas fuentes de dicha energía eran la radiación solar y las descargas eléctricas. [1] Utilizando una gran cantidad de energía metabólica y la enzima nitrogenasa , algunas bacterias y cianobacterias convierten el N 2 atmosférico en NH 3 , un proceso conocido como fijación biológica de nitrógeno (FBN). [4] El análogo antropogénico de la FBN es el proceso Haber-Bosch , en el que el H 2 reacciona con el N 2 atmosférico a altas temperaturas y presiones para producir NH 3 . [5] Por último, el N 2 se convierte en NO mediante la energía de los rayos , que es insignificante en los ecosistemas templados actuales, o mediante la combustión de combustibles fósiles . [1]
Hasta 1850, la FBN natural, la FBN inducida por el cultivo (por ejemplo, la plantación de cultivos leguminosos ) y la materia orgánica incorporada eran las únicas fuentes de N para la producción agrícola. [5] Cerca del cambio de siglo, el Nr de los depósitos de guano y nitrato de sodio se cosechaba y exportaba desde las áridas islas del Pacífico y los desiertos sudamericanos. [5] A fines de la década de 1920, los primeros procesos industriales, aunque ineficientes, se usaban comúnmente para producir NH 3 . [1] Debido a los esfuerzos de Fritz Haber y Carl Bosch , el proceso Haber-Bosch se convirtió en la mayor fuente de fertilizante nitrogenado después de la década de 1950 y reemplazó a la FBN como la fuente dominante de producción de NH 3 . [5] De 1890 a 1990, el Nr creado antropogénicamente aumentó casi nueve veces. [1] Durante este tiempo, la población humana se triplicó, en parte debido al aumento de la producción de alimentos.
Desde la Revolución Industrial , una fuente adicional de aporte antropogénico de N ha sido la combustión de combustibles fósiles , que se utiliza para liberar energía (por ejemplo, para impulsar automóviles). A medida que se queman combustibles fósiles , las altas temperaturas y presiones proporcionan energía para producir NO a partir de la oxidación de N 2 . [1] Además, cuando se extrae y se quema combustible fósil , el N fósil puede volverse reactivo (es decir, emisiones de NO x ). [1] Durante la década de 1970, los científicos comenzaron a reconocer que los aportes de N se estaban acumulando en el medio ambiente y afectando a los ecosistemas. [1]
Impactos de los aportes antropogénicos en el ciclo del nitrógeno
Entre 1600 y 1990, la creación global de nitrógeno reactivo (Nr) había aumentado casi un 50%. [6] Durante este período, las emisiones atmosféricas de especies de Nr aumentaron un 250% y la deposición en ecosistemas marinos y terrestres aumentó más del 200%. [6] Además, se informó de un aumento de cuatro veces en los flujos de N inorgánico disuelto de los ríos hacia las costas. [6] El nitrógeno es un nutriente limitante crítico en muchos sistemas, incluidos los bosques, los humedales y los ecosistemas costeros y marinos; por lo tanto, este cambio en las emisiones y la distribución de Nr ha tenido consecuencias sustanciales para los ecosistemas acuáticos y terrestres. [7] [8]
Atmósfera
| Tipos de alimentos | Emisiones de gases de efecto invernadero (g CO 2 -C eq por g de proteína) |
|---|---|
| Carne de rumiantes | |
| Acuicultura de recirculación | |
| Pesca de arrastre | |
| Acuicultura sin recirculación | |
| Cerdo | |
| Aves de corral | |
| Lácteos | |
| Pesca no basada en el arrastre | |
| Huevos | |
| Raíces almidonadas | |
| Trigo | |
| Maíz | |
| Legumbres |
Las entradas de N atmosférico incluyen principalmente óxidos de N (NO x ), amoníaco (NH 3 ) y óxido nitroso (N 2 O) de los ecosistemas acuáticos y terrestres, [4] y NO x de la combustión de combustibles fósiles y biomasa. [1]
En los agroecosistemas , la aplicación de fertilizantes ha aumentado la nitrificación microbiana (proceso aeróbico en el que los microorganismos oxidan el amonio [NH 4 + ] a nitrato [NO 3 − ]) y la desnitrificación (proceso anaeróbico en el que los microorganismos reducen el NO 3 − al gas nitrógeno atmosférico [N 2 ]). Ambos procesos liberan naturalmente óxido nítrico (NO) y óxido nitroso (N 2 O) a la atmósfera. [4] De particular preocupación es el N 2 O, que tiene una vida atmosférica promedio de 114-120 años, [10] y es 300 veces más efectivo que el CO 2 como gas de efecto invernadero . [4] El NO x producido por procesos industriales, automóviles y fertilización agrícola y el NH 3 emitido por los suelos (es decir, como un subproducto adicional de la nitrificación) [4] y las operaciones ganaderas se transportan a ecosistemas a sotavento, lo que influye en el ciclo del N y las pérdidas de nutrientes. Se han citado seis efectos principales de las emisiones de NO x y NH 3 : [1] 1) disminución de la visibilidad atmosférica debido a los aerosoles de amonio ( materia particulada fina [PM]); 2) concentraciones elevadas de ozono ; 3) el ozono y el PM afectan la salud humana (por ejemplo, enfermedades respiratorias , cáncer ); 4) aumentos en el forzamiento radiativo y el cambio climático global ; 5) disminución de la productividad agrícola debido a la deposición de ozono ; y 6) acidificación de los ecosistemas [11] y eutrofización .
Biosfera
| Parte de una serie sobre |
| Ciclos biogeoquímicos |
|---|
 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Los ecosistemas terrestres y acuáticos reciben aportes de Nr de la atmósfera a través de la deposición húmeda y seca. [1] Las especies atmosféricas de Nr pueden depositarse en los ecosistemas en las precipitaciones (por ejemplo, NO 3 − , NH 4 + , compuestos orgánicos de N), como gases (por ejemplo, NH 3 y ácido nítrico gaseoso [HNO 3 ]), o como aerosoles (por ejemplo, nitrato de amonio [NH 4 NO 3 ]). [1] Los ecosistemas acuáticos reciben nitrógeno adicional de la escorrentía superficial y de los aportes fluviales . [8]
El aumento de la deposición de N puede acidificar los suelos, arroyos y lagos y alterar la productividad de los bosques y pastizales. En los ecosistemas de pastizales, los aportes de N han producido aumentos iniciales en la productividad seguidos de disminuciones a medida que se superan los umbrales críticos. [1] También se han demostrado los efectos del nitrógeno en la biodiversidad , el ciclo del carbono y los cambios en la composición de las especies . En áreas altamente desarrolladas de sistemas costeros oceánicos y estuarinos cercanos a la costa, los ríos aportan aportes de N directos (p. ej., escorrentía superficial ) e indirectos (p. ej., contaminación de las aguas subterráneas) de los agroecosistemas. [8] El aumento de los aportes de N puede provocar la acidificación del agua dulce y la eutrofización de las aguas marinas.
Ecosistemas terrestres
Impactos en la productividad y el ciclo de nutrientes
Gran parte del crecimiento terrestre en los sistemas templados está limitado por el N; por lo tanto, los aportes de N (es decir, a través de la deposición y la fertilización) pueden aumentar la disponibilidad de N, lo que aumenta temporalmente la absorción de N, el crecimiento vegetal y microbiano, y la acumulación de N en la biomasa vegetal y la materia orgánica del suelo . [12] La incorporación de mayores cantidades de N en la materia orgánica disminuye las relaciones C:N, aumentando la liberación de N mineral (NH 4 + ) durante la descomposición de la materia orgánica por microbios heterotróficos (es decir, amonificación ). [13] A medida que aumenta la amonificación, también lo hace la nitrificación del N mineralizado. Debido a que la nitrificación y la desnitrificación microbianas son "permeables", se espera que la deposición de N aumente las emisiones de gases traza. [14] Además, con el aumento de la acumulación de NH 4 + en el suelo, los procesos de nitrificación liberan iones de hidrógeno, que acidifican el suelo. NO 3 − , el producto de la nitrificación, es muy móvil y puede lixiviarse del suelo, junto con minerales alcalinos cargados positivamente como el calcio y el magnesio. [4] En suelos ácidos, los iones de aluminio movilizados pueden alcanzar concentraciones tóxicas, afectando negativamente tanto a los ecosistemas terrestres como a los acuáticos adyacentes.
Las fuentes antropogénicas de N generalmente llegan a los bosques de las tierras altas a través de la deposición . [15] Una posible preocupación del aumento de la deposición de N debido a las actividades humanas es la alteración del ciclo de nutrientes en los ecosistemas forestales. Numerosos estudios han demostrado impactos tanto positivos como negativos de la deposición atmosférica de N en la productividad forestal y el almacenamiento de carbono. El N añadido suele ser inmovilizado rápidamente por microbios , [16] y el efecto del N disponible restante depende de la capacidad de la comunidad vegetal para la absorción de N. [17] En sistemas con alta absorción, el N se asimila en la biomasa vegetal, lo que conduce a una mayor productividad primaria neta (PPN) y posiblemente a un mayor secuestro de carbono a través de una mayor capacidad fotosintética. Sin embargo, las respuestas del ecosistema a las adiciones de N dependen de muchos factores específicos del sitio, incluidos el clima, el historial de uso de la tierra y la cantidad de adiciones de N. Por ejemplo, en el noreste de los Estados Unidos, las masas de madera dura que reciben aportes crónicos de N han demostrado una mayor capacidad para retener N y aumentar la productividad primaria neta anual (PPNA) que las masas de coníferas. [18] Una vez que la entrada de N excede la demanda del sistema, el N puede perderse a través de la lixiviación y los flujos de gas. Cuando el N disponible excede la capacidad de absorción del ecosistema (es decir, vegetación, suelo y microbios , etc.), se produce la saturación de N y el exceso de N se pierde en las aguas superficiales, subterráneas y en la atmósfera. [12] [17] [18] La saturación de N puede provocar desequilibrios de nutrientes (por ejemplo, pérdida de calcio debido a la lixiviación de nitratos) y posible declive forestal. [13]
Un estudio de 15 años sobre las adiciones crónicas de N en el programa de Investigación Ecológica a Largo Plazo ( LTER ) de Harvard Forest ha dilucidado muchos de los impactos del aumento de la deposición de nitrógeno en el ciclo de nutrientes en los bosques templados. Encontró que las adiciones crónicas de N resultaron en mayores pérdidas por lixiviación, mayor mortalidad de pinos y cese de la acumulación de biomasa. [18] Otro estudio informó que las adiciones crónicas de N resultaron en la acumulación de N no fotosintético y posteriormente redujeron la capacidad fotosintética, lo que supuestamente conduce a un estrés de carbono severo y mortalidad. [17] Estos hallazgos niegan las hipótesis anteriores de que el aumento de las entradas de N aumentaría la PPN y el secuestro de carbono .
Impactos en la diversidad de especies vegetales
Muchas comunidades vegetales han evolucionado en condiciones de bajos nutrientes; por lo tanto, el aumento de los aportes de N puede alterar las interacciones bióticas y abióticas, lo que lleva a cambios en la composición de la comunidad. Varios estudios de adición de nutrientes han demostrado que el aumento de los aportes de N conduce al predominio de especies vegetales de rápido crecimiento, con disminuciones asociadas en la riqueza de especies. [19] [20] [21] Las especies de rápido crecimiento tienen una mayor afinidad por la absorción de nitrógeno y desplazarán a las especies vegetales de crecimiento más lento al bloquear el acceso a la luz solar con su mayor biomasa sobre el suelo. [22] Otros estudios han encontrado que las respuestas secundarias del sistema al enriquecimiento de N, incluida la acidificación del suelo y los cambios en las comunidades micorrízicas , han permitido que las especies tolerantes al estrés superen a las especies sensibles. [11] [23] Los árboles que tienen asociaciones micorrízicas arbusculares tienen más probabilidades de beneficiarse de un aumento en el nitrógeno del suelo, ya que estos hongos son incapaces de descomponer el nitrógeno orgánico del suelo. [24] Otros dos estudios encontraron evidencia de que el aumento de la disponibilidad de N ha resultado en disminuciones en brezales con diversidad de especies . Los brezales se caracterizan por tener suelos pobres en N, que excluyen los pastos que lo demandan; sin embargo, con el aumento de la deposición de N y la acidificación del suelo , los pastizales invasores reemplazan a los brezales de tierras bajas. [25] [26]
En un estudio experimental más reciente de fertilización con N y perturbación (es decir, labranza) en sucesiones de campos antiguos, se encontró que la riqueza de especies disminuyó con el aumento de N, independientemente del nivel de perturbación. Los experimentos de competencia mostraron que los dominantes competitivos excluían a las especies competitivamente inferiores entre eventos de perturbación. Con mayores aportes de N, la competencia pasó del subterráneo al superficial (es decir, competencia por luz), y las tasas de colonización de parches disminuyeron significativamente. Estos cambios internos pueden afectar dramáticamente a la comunidad al cambiar el equilibrio de las compensaciones entre competencia y colonización entre especies. [21] En sistemas basados en parches, la coexistencia regional puede ocurrir a través de compensaciones en las habilidades competitivas y colonizadoras dadas tasas de perturbación suficientemente altas. [27] Es decir, con una clasificación inversa de las habilidades competitivas y colonizadoras, las plantas pueden coexistir en el espacio y el tiempo a medida que la perturbación elimina a los competidores superiores de los parches, lo que permite el establecimiento de colonizadores superiores. Sin embargo, como demostraron Wilson y Tilman, el aumento de los aportes de nutrientes puede anular las compensaciones, lo que resulta en la exclusión competitiva de estos colonizadores superiores/competidores pobres. [21]
Ecosistemas acuáticos
Los ecosistemas acuáticos también muestran respuestas variadas al enriquecimiento de nitrógeno. La carga de NO 3 − de los ecosistemas terrestres saturados de N puede provocar la acidificación de los sistemas de agua dulce aguas abajo y la eutrofización de los sistemas marinos aguas abajo. La acidificación del agua dulce puede causar toxicidad por aluminio y mortalidad de especies de peces sensibles al pH. Debido a que los sistemas marinos generalmente tienen un contenido limitado de nitrógeno, los aportes excesivos de N pueden provocar la degradación de la calidad del agua debido a floraciones de algas tóxicas, deficiencia de oxígeno, pérdida de hábitat, disminución de la biodiversidad y pérdidas de pesca. [8]
Acidificación de las aguas dulces
La deposición atmosférica de N en paisajes terrestres puede transformarse a través de procesos microbianos del suelo en nitrógeno biológicamente disponible, lo que puede provocar la acidificación de las aguas superficiales y la pérdida de biodiversidad . Los aportes de NO 3 − y NH 4 + de los sistemas terrestres y la atmósfera pueden acidificar los sistemas de agua dulce cuando hay poca capacidad de amortiguación debido a la acidificación del suelo . [8] La contaminación por N en Europa, el noreste de los Estados Unidos y Asia es una preocupación actual por la acidificación del agua dulce . [28] Los estudios de acidificación de lagos en el Área Experimental del Lago (ELA) en el noroeste de Ontario demostraron claramente los efectos negativos del aumento de la acidez en una especie de pez nativa: el reclutamiento y el crecimiento de la trucha de lago (Salvelinus namaycush) disminuyeron drásticamente debido a la extirpación de sus principales especies de presas durante la acidificación. [29] El nitrógeno reactivo de la agricultura, la cría de animales, los fertilizantes, los sistemas sépticos y otras fuentes han aumentado las concentraciones de nitrato en las vías fluviales de la mayoría de las naciones industrializadas. En menos de una década, las concentraciones de nitratos en 1.000 lagos noruegos se duplicaron. En el noreste de Estados Unidos y la mayor parte de Europa, la concentración de nitratos se multiplicó por diez o quince en el último siglo. El nitrógeno reactivo puede contaminar el agua potable al verterlo en arroyos, lagos, ríos y aguas subterráneas. Sólo en Estados Unidos, hasta un 20% de las fuentes de aguas subterráneas superan el límite de concentración de nitratos en el agua potable establecido por la Organización Mundial de la Salud. Estas altas concentraciones pueden provocar la "enfermedad del bebé azul", en la que los iones de nitrato debilitan la capacidad de la sangre para transportar oxígeno. Los estudios también han vinculado las altas concentraciones de nitratos con problemas reproductivos y la propensión a algunos tipos de cáncer, como el de vejiga y el de ovario. [30]
Eutrofización de los sistemas marinos
La urbanización, la deforestación y las actividades agrícolas contribuyen en gran medida a los aportes de sedimentos y nutrientes a las aguas costeras a través de los ríos. [8] El aumento de los aportes de nutrientes a los sistemas marinos ha mostrado aumentos a corto plazo en la productividad y los rendimientos pesqueros, y efectos perjudiciales a largo plazo de la eutrofización . La triplicación de las cargas de NO 3 − en el río Misisipi en la segunda mitad del siglo XX se ha correlacionado con el aumento de los rendimientos pesqueros en las aguas que rodean el delta del Misisipi; [31] sin embargo, estos aportes de nutrientes han producido hipoxia estacional (concentraciones de oxígeno inferiores a 2-3 mg L −1 , " zonas muertas ") en el Golfo de México . [1] [8] En los sistemas estuarinos y costeros, los altos aportes de nutrientes aumentan la producción primaria (p. ej., fitoplancton , pastos marinos, macroalgas), que aumentan la turbidez con las consiguientes disminuciones en la penetración de la luz en toda la columna de agua. En consecuencia, el crecimiento de la vegetación sumergida disminuye, lo que reduce la complejidad del hábitat y la producción de oxígeno. El aumento de la producción primaria (es decir, fitoplancton, macroalgas, etc.) conduce a un flujo de carbono hacia las aguas del fondo cuando la materia orgánica en descomposición (es decir, la producción primaria senescente) se hunde y es consumida por bacterias aeróbicas en la parte inferior de la columna de agua. Como resultado, el consumo de oxígeno en las aguas del fondo es mayor que la difusión de oxígeno desde las aguas superficiales. Además, ciertas floraciones de algas denominadas floraciones de algas nocivas (FAN) producen toxinas que pueden actuar como compuestos neuromusculares o dañinos para los órganos. Estas floraciones de algas pueden ser dañinas para otras formas de vida marina, así como para los seres humanos. [32] [33]
Integración
Las respuestas del sistema antes mencionadas a los aportes de nitrógeno reactivo (Nr) se estudian casi exclusivamente por separado; sin embargo, la investigación indica cada vez más que los problemas de carga de nitrógeno están vinculados por múltiples vías que transportan nutrientes a través de los límites del sistema. [1] Esta transferencia secuencial entre ecosistemas se denomina cascada de nitrógeno. [6] (ver ilustración del Programa de las Naciones Unidas para el Medio Ambiente). Durante la cascada, algunos sistemas acumulan Nr, lo que da lugar a un desfase temporal en la cascada y a mayores efectos del Nr en el entorno en el que se acumula. En última instancia, los aportes antropogénicos de Nr se acumulan o se desnitrifican; sin embargo, se ha avanzado poco en la determinación de la importancia relativa de la acumulación y la desnitrificación de Nr , lo que se ha debido principalmente a una falta de integración entre las disciplinas científicas. [1] [34]
La mayor parte del Nr aplicado a los agroecosistemas globales se transmite en cascada a través de la atmósfera y los ecosistemas acuáticos y terrestres hasta que se convierte en N 2 , principalmente a través de la desnitrificación . [1] Aunque la desnitrificación terrestre produce intermediarios gaseosos (óxido nítrico [NO] y óxido nitroso [N 2 O]), el último paso (la producción microbiana de N 2 ) es fundamental porque el N 2 atmosférico es un sumidero de Nr. [34] Muchos estudios han demostrado claramente que las franjas de amortiguación y los humedales gestionados pueden eliminar cantidades significativas de nitrato (NO 3 − ) de los sistemas agrícolas a través de la desnitrificación . [35] Tal gestión puede ayudar a atenuar los efectos en cascada indeseables y eliminar la acumulación de Nr ambiental. [1]
Las actividades humanas dominan los ciclos de N globales y la mayoría de las regiones. [36] Las entradas de N han mostrado consecuencias negativas tanto para el ciclo de nutrientes como para la diversidad de especies nativas en sistemas terrestres y acuáticos. De hecho, debido a los impactos a largo plazo en las redes alimentarias, las entradas de Nr se consideran ampliamente el problema de contaminación más crítico en los sistemas marinos. [8] En los ecosistemas terrestres y acuáticos, las respuestas al enriquecimiento de N varían; sin embargo, un tema general recurrente es la importancia de los umbrales (por ejemplo, la saturación de nitrógeno ) en la capacidad de retención de nutrientes del sistema. Para controlar la cascada de N, debe haber una integración de disciplinas científicas y más trabajo sobre el almacenamiento de Nr y las tasas de desnitrificación . [34]
Véase también
Referencias
- ^ abcdefghijklmnopqrst Galloway, JN; Aber, JD; Erisman, JNW; Seitzinger, SP; Howarth, RW; Cowling, EB; Cosby, BJ (2003). "La cascada del nitrógeno". BioScience . 53 (4): 341. doi : 10.1641/0006-3568(2003)053[0341:TNC]2.0.CO;2 . S2CID 3356400.
- ^ ab Alley et al. 2007. IPCC Climate Change 2007: The Physical Science Basis. Contribución del Grupo de trabajo I al Tercer informe de evaluación del Grupo Intergubernamental de Expertos sobre el Cambio Climático. Resumen del informe para los responsables de las políticas (SPM) Archivado el 16 de julio de 2011 en Wayback Machine .
- ^ Nemecek, T.; Poore, J. (1 de junio de 2018). "Reducción de los impactos ambientales de los alimentos a través de productores y consumidores". Science . 360 (6392): 987–992. Bibcode :2018Sci...360..987P. doi : 10.1126/science.aaq0216 . ISSN 0036-8075. PMID 29853680. S2CID 206664954.
- ^ abcdef Schlesinger, WH 1997. Biogeoquímica: Un análisis del cambio global , San Diego, CA.
- ^ abcd Smil, V. 2001. Enriqueciendo la tierra: Fritz Haber, Carl Bosch y la transformación de la producción mundial de alimentos . MIT Press, Cambridge, MA.
- ^ abcd Galloway, James N.; Cowling, Ellis B. (2002). "Nitrógeno reactivo y el mundo: 200 años de cambio". Ambio: A Journal of the Human Environment . 31 (2): 64–71. Bibcode :2002Ambio..31...64G. doi :10.1579/0044-7447-31.2.64. PMID 12078011. S2CID 8104525.
- ^ Vitousek, P.; Howarth, R. (1991). "Limitación de nitrógeno en la tierra y en el mar: ¿cómo puede ocurrir?". Biogeoquímica . 13 (2). doi :10.1007/BF00002772. S2CID 93106377.
- ^ abcdefgh Rabalais, Nancy N. (2002). "Nitrógeno en ecosistemas acuáticos". Ambio: A Journal of the Human Environment . 31 (2): 102–12. doi :10.1639/0044-7447(2002)031[0102:NIAE]2.0.CO;2. PMID 12077998.
- ^ Michael Clark; Tilman, David (noviembre de 2014). "Las dietas globales vinculan la sostenibilidad ambiental y la salud humana". Nature . 515 (7528): 518–522. Bibcode :2014Natur.515..518T. doi :10.1038/nature13959. ISSN 1476-4687. PMID 25383533. S2CID 4453972.
- ^ John T. Houghton , Y. Ding, DJ Griggs, M. Noguer, PJ van der Linden, X. Dai, K. Maskell y CA Johnson. 2001. IPCC Climate Change 2001: The Scientific Basis. Contribución del Grupo de Trabajo I al Tercer Informe de Evaluación del Grupo Intergubernamental de Expertos sobre el Cambio Climático . Cambridge University Press]
- ^ ab Houdijk, ALFM; Verbeek, PJM; Dijk, HFG; Roelofs, JGM (1993). "Distribución y declive de especies de brezales herbáceos en peligro de extinción en relación con la composición química del suelo". Plantas y suelos . 148 (1): 137–143. Bibcode :1993PlSoi.148..137H. doi :10.1007/BF02185393. S2CID 22600629.
- ^ ab Aber, JD, KJ Nadelhoffer, P. Steudler y JM Melillo. 1989. "Saturación de nitrógeno en ecosistemas forestales del norte". Biociencia 39:378–386
- ^ ab Aber, JD (1992). "Ciclado del nitrógeno y saturación de nitrógeno en ecosistemas forestales templados". Tendencias en ecología y evolución . 7 (7): 220–224. doi :10.1016/0169-5347(92)90048-G. PMID 21236013.
- ^ Matson, P; Lohse, KA; Hall, SJ (2002). "La globalización de la deposición de nitrógeno: consecuencias para los ecosistemas terrestres". Ambio . 31 (2): 113–9. doi :10.1639/0044-7447(2002)031[0113:tgondc]2.0.co;2. JSTOR 4315223. PMID 12077999.
- ^ Aber, John D.; Goodale, Christine L .; Ollinger, Scott V.; Smith, Marie-Louise; Magill, Alison H.; Martin, Mary E.; Hallett, Richard A.; Stoddard, John L. (2003). "¿La deposición de nitrógeno está alterando el estado del nitrógeno de los bosques del noreste?". BioScience . 53 (4): 375. doi : 10.1641/0006-3568(2003)053[0375:INDATN]2.0.CO;2 . JSTOR 1314369. S2CID 54037020.
- ^ Nadelhoffer, KJ; Downs, MR; Fry, B. (1999). "Sumideros para adiciones enriquecidas con 15N a un bosque de robles y una plantación de pino rojo". Aplicaciones ecológicas . 9 : 72–86. doi :10.1890/1051-0761(1999)009[0072:SFNEAT]2.0.CO;2.
- ^ abc Bauer, GA; Bazzaz, FA; Minocha, R.; Long, S.; Magill, A.; Aber, J.; Berntson, GM (2004). "Efectos de las adiciones crónicas de N en la química de los tejidos, la capacidad fotosintética y el potencial de secuestro de carbono de una población de pino rojo (Pinus resinosa Ait.) en el noreste de los Estados Unidos". Ecología y gestión forestal . 196 : 173–186. doi :10.1016/j.foreco.2004.03.032.
- ^ abc Magill, AH; Aber, JD; Currie, WS; Nadelhoffer, KJ; Martin, ME; McDowell, WH; Melillo, JM; Steudler, P. (2004). "Respuesta del ecosistema a 15 años de adiciones crónicas de nitrógeno en el LTER de Harvard Forest, Massachusetts, EE. UU." Ecología y gestión forestal . 196 : 7–28. doi :10.1016/j.foreco.2004.03.033.
- ^ Laura Foster Huenneke; Steven P. Hamburg; Roger Koide; Harold A. Mooney; Peter M. Vitousek (1990). "Efectos de los recursos del suelo en la invasión de plantas y la estructura de la comunidad en los pastizales serpentinos de California". Ecología . 71 (2): 478–491. Bibcode :1990Ecol...71..478H. doi :10.2307/1940302. JSTOR 1940302.
- ^ Tilman, D. (1997). "Invasibilidad de la comunidad, limitación del reclutamiento y biodiversidad de pastizales". Ecology . 78 : 81–83. doi :10.1890/0012-9658(1997)078[0081:CIRLAG]2.0.CO;2.
- ^ abc Wilson, SD; Tilman, D. (2002). "Variación cuadrática en la riqueza de especies de campos antiguos a lo largo de gradientes de perturbación y nitrógeno". Ecología . 83 (2): 492. doi :10.1890/0012-9658(2002)083[0492:QVIOFS]2.0.CO;2.
- ^ Wamelink, GWW; van Dobben, HF; Mol-Dijkstra, JP; Schouwenberg, EPAG; Kros, J.; de Vries, W.; Berendse, F. (septiembre de 2009). "Efecto de la reducción de la deposición de nitrógeno sobre la biodiversidad y el secuestro de carbono". Ecología y Gestión Forestal . 258 (8): 1774-1779. doi :10.1016/j.foreco.2008.10.024.
- ^ Egerton-Warburton, LM; Allen, EB (2000). "Cambios en las comunidades micorrízicas arbusculares a lo largo de un gradiente de deposición de nitrógeno antropogénico". Aplicaciones ecológicas . 10 (2): 484. doi :10.1890/1051-0761(2000)010[0484:SIAMCA]2.0.CO;2.
- ^ Quinn Thomas, R.; Canham, Charles D.; Weathers, Kathleen C.; Goodale, Christine L. (22 de diciembre de 2009). "Aumento del almacenamiento de carbono en los árboles en respuesta a la deposición de nitrógeno en los EE. UU." Nature Geoscience . 3 (1): 13–17. doi :10.1038/ngeo721. ISSN 1752-0908.
- ^ Aerts, Rien; Berendse, Frank (agosto de 1988). "El efecto del aumento de la disponibilidad de nutrientes en la dinámica de la vegetación en brezales húmedos". Vegetatio . 76 (1–2): 63–69. doi :10.1007/BF00047389. ISSN 0042-3106. S2CID 34882407.
- ^ Bobbink, R.; Heil, GW; Raessen, MB (1992). "Deposición atmosférica y procesos de intercambio de dosel en ecosistemas de brezales". Contaminación ambiental . 75 (1): 29–37. Bibcode :1992EPoll..75...29B. doi :10.1016/0269-7491(92)90053-D. PMID 15092046.
- ^ Hastings, A. (1980). "Perturbación, coexistencia, historia y competencia por el espacio". Biología teórica de poblaciones . 18 (3): 363–373. doi :10.1016/0040-5809(80)90059-3.
- ^ Driscoll, CT, GB Lawrence, AJ Bulger, TJ Butler, CS Cronan, C. Eagar, KF Lambert, GE Likens, JL Stoddard y KC Weathers. 2001. Deposición ácida en el noreste de Estados Unidos: fuentes y aportes, efectos en los ecosistemas y estrategias de gestión. pp. 180-198
- ^ Mills, KH; Chalanchuk, SM; Allan, DJ (2000). "Recuperación de las poblaciones de peces en el lago 223 a partir de la acidificación experimental". Revista Canadiense de Ciencias Pesqueras y Acuáticas . 57 : 192–204. doi :10.1139/f99-186.
- ^ Fields, Scott (julio de 2004). "Nitrógeno global: ciclo fuera de control". Environmental Health Perspectives . 112 (10): A556–A563. doi :10.1289/ehp.112-a556. PMC 1247398 . PMID 15238298.
- ^ Grimes, Churchill B. (2001). "Producción pesquera y descarga del río Mississippi". Pesca . 26 (8): 17–26. Bibcode :2001Fish...26g..17G. doi :10.1577/1548-8446(2001)026<0017:FPATMR>2.0.CO;2.
- ^ Skulberg, Olav M. ; Codd, Geoffrey A.; Carmichael, Wayne W. (1 de enero de 1984). "Floraciones tóxicas de algas verdeazuladas en Europa: un problema creciente". Ambio . 13 (4): 244–247. JSTOR 4313034.
- ^ Smith, VH; Tilman, GD; Nekola, JC (1999). "Eutrofización: impactos del exceso de aportes de nutrientes en los ecosistemas de agua dulce, marinos y terrestres". Contaminación ambiental . 100 (1–3): 179–196. doi :10.1016/s0269-7491(99)00091-3. PMID 15093117. S2CID 969039.
- ^ abc Davidson, EA; Seitzinger, S. (2006). "El enigma del progreso en la investigación sobre desnitrificación". Aplicaciones ecológicas . 16 (6): 2057–2063. doi :10.1890/1051-0761(2006)016[2057:TEOPID]2.0.CO;2. PMID 17205889.
- ^ Jackson, RD; Allen-Diaz, B.; Oates, LG; Tate, KW (2006). "Nitrato de agua de manantial aumentado con la eliminación del pastoreo de ganado en una sabana de robles de California". Ecosystems . 9 (2): 254. Bibcode :2006Ecosy...9..254J. doi :10.1007/s10021-005-0166-7. S2CID 24450808.
- ^ Galloway, JN; Dentener, FJ; Capone, DG; Boyer, EW; Howarth, RW; Seitzinger, SP; Asner, GP; Cleveland, CC; Green, PA; Holland, EA; Karl, DM; Michaels, AF; Porter, JH; Townsend, AR; Vöosmarty, CJ (2004). "Ciclos del nitrógeno: pasado, presente y futuro" (PDF) . Biogeoquímica . 70 (2): 153. Bibcode :2004Biogc..70..153G. doi :10.1007/s10533-004-0370-0. JSTOR 4151466. S2CID 98109580. Archivado desde el original (PDF) el 2005-11-11.
Lectura adicional
- Good, AG; Beatty, PH (2011). "Fertilizar la naturaleza: una tragedia de excesos en los bienes comunes". PLOS Biology . 9 (8): e1001124. doi : 10.1371/journal.pbio.1001124 . PMC 3156687 . PMID 21857803..
- Scarsbrook M.; Barquin J.; Gray D. (2007). Manantiales de agua fría de Nueva Zelanda y su biodiversidad (PDF) . ISBN 978-0-478-14289-1. ISSN 1173-2946.
{{cite book}}:|journal=ignorado ( ayuda ) - Olde Venterink, H.; Wassen, MJ; Verkroost, AWM; De Ruiter, PC (2003). "Species Richness–Productivity Patterns Differ Between N-, P-, and K-Limited Wetlands" (PDF) . Ecología . 84 (8): 2191–2199. Bibcode :2003Ecol...84.2191O. doi :10.1890/01-0639. JSTOR 3450042. Archivado desde el original (PDF) el 2016-03-03 . Consultado el 2009-09-03 .
