Gasterópodos
| Gasterópodos Rango temporal: | |
|---|---|
 | |
| Varios gasterópodos de diferentes tipos: babosa negra (una babosa), Haliotis asinina (una abulón), Cornu aspersum (un caracol terrestre), Notarchus indicus (una liebre de mar), Patella vulgata (una lapa) y Polycera aurantiomarginata (un nudibranquio). | |
| Clasificación científica | |
| Dominio: | Eucariota |
| Reino: | Animalia |
| Filo: | Moluscos |
| Clase: | Gasterópodos Cuvier , 1795 [2] |
| Subclases | |
| Diversidad [3] [4] | |
| 65.000 a 80.000 especies | |
| Sinónimos [5] | |
| |
Los gasterópodos ( / ˈ ɡ æ s t r ə p ɒ d z / ), comúnmente conocidos como babosas y caracoles , pertenecen a una gran clase taxonómica de invertebrados dentro del filo Mollusca llamado Gastropoda ( / ɡ æ s ˈ t r ɒ p ə d ə / ). [5]
Esta clase comprende caracoles y babosas de agua salada, de agua dulce y de tierra. Existen miles de especies de caracoles marinos y babosas , así como caracoles de agua dulce , lapas de agua dulce , caracoles terrestres y babosas .
La clase Gastropoda es una clase diversa y muy exitosa de moluscos dentro del filo Mollusca. Contiene una gran cantidad de especies nombradas, superada solo por los insectos en número total. La historia fósil de esta clase se remonta al Cámbrico tardío . En 2017 , se conocen [actualizar]721 familias de gasterópodos, de las cuales 245 están extintas y aparecen solo en el registro fósil , mientras que 476 están actualmente existentes con o sin registro fósil. [6]
Los gasterópodos (antes conocidos como univalvos y a veces escritos "Gasteropoda") son una parte importante del filo Mollusca, y son la clase más diversificada del filo , con 65.000 a 80.000 [3] [4] especies vivas de caracoles y babosas . La anatomía , el comportamiento, la alimentación y las adaptaciones reproductivas de los gasterópodos varían significativamente de un clado o grupo a otro, por lo que es difícil establecer muchas generalidades para todos los gasterópodos.
La clase Gastropoda presenta una extraordinaria diversificación de hábitats . Sus representantes viven en jardines, bosques, desiertos y montañas; en pequeñas zanjas, grandes ríos y lagos; en estuarios , marismas , zonas intermareales rocosas , zonas submareales arenosas, profundidades abisales de los océanos, incluidas las fuentes hidrotermales , y en numerosos otros nichos ecológicos, incluidos los parásitos .
Aunque el nombre "caracol" puede aplicarse, y a menudo se aplica, a todos los miembros de esta clase, comúnmente esta palabra se refiere sólo a aquellas especies con una concha externa lo suficientemente grande como para que las partes blandas puedan retraerse completamente en ella. Las babosas son gasterópodos que no tienen concha o tienen una concha interna muy pequeña; las semibabosas son gasterópodos que tienen una concha en la que pueden retraerse parcialmente, pero no por completo.
Las especies marinas con concha de gasterópodos incluyen especies como abulón , caracoles marinos , caracoles marinos , caracoles marinos de mar y muchos otros caracoles marinos que producen conchas enrolladas en la etapa adulta, aunque en algunos casos, la espiral puede no ser muy visible, por ejemplo, en los cauris . En varias familias de especies, como todas las diversas lapas , la concha está enrollada solo en la etapa larvaria y, a partir de entonces, es una estructura cónica simple.
Etimología
En la literatura científica, los gasterópodos fueron descritos como "gasterópodos" por Georges Cuvier en 1795. [2] La palabra gasterópodo proviene del griego γαστήρ ( gastḗr 'estómago') y πούς ( poús 'pie'), una referencia al hecho de que el "pie" del animal está ubicado debajo de sus entrañas. [7]
El nombre anterior "univalvo" significa una válvula (o concha), en contraste con los bivalvos , como las almejas, que tienen dos valvas o conchas.
Diversidad
En todos los niveles taxonómicos , los gasterópodos ocupan el segundo lugar, después de los insectos, en términos de diversidad . [8]
Los gasterópodos son los moluscos con mayor número de especies nombradas. Sin embargo, las estimaciones del número total de especies de gasterópodos varían ampliamente, dependiendo de las fuentes citadas. El número de especies de gasterópodos se puede determinar a partir de las estimaciones del número de especies descritas de moluscos con nombres aceptados: alrededor de 85.000 (mínimo 50.000, máximo 120.000). [9] Pero una estimación del número total de moluscos, incluidas las especies no descritas, es de alrededor de 240.000 especies. [10] La estimación de 85.000 moluscos incluye 24.000 especies descritas de gasterópodos terrestres. [9]
Según distintas estimaciones (basadas en distintas fuentes) para los gasterópodos acuáticos, hay unas 30.000 especies de gasterópodos marinos y unas 5.000 especies de gasterópodos de agua dulce y salobre . Aún quedan muchas especies de aguas profundas por descubrir, ya que solo se ha estudiado biológicamente el 0,0001% del fondo marino. [11] [12] El número total de especies vivas de caracoles de agua dulce es de unas 4.000. [13]
Las especies de gasterópodos recientemente extintas (extintas desde 1500) suman 444, 18 especies están ahora extintas en estado salvaje (pero aún existen en cautiverio) y 69 especies están "posiblemente extintas". [14]
El número de especies prehistóricas (fósiles) de gasterópodos es de al menos 15.000 especies. [15]
En los hábitats marinos, el talud continental y la vertiente continental albergan la mayor diversidad, mientras que la plataforma continental y las profundidades abisales tienen una baja diversidad de gasterópodos marinos. [16]
Hábitat

Los gasterópodos se encuentran en una amplia gama de hábitats acuáticos y terrestres, desde fosas oceánicas profundas hasta desiertos.
Algunos de los gasterópodos más conocidos son los gasterópodos terrestres (los caracoles y las babosas). Algunos viven en agua dulce, pero la mayoría de las especies de gasterópodos que se conocen viven en un entorno marino.
Los gasterópodos tienen una distribución mundial, desde las zonas cercanas al Ártico y la Antártida hasta los trópicos. Se han adaptado a casi todas las formas de vida en la Tierra, habiendo colonizado casi todos los medios disponibles.
En hábitats donde no hay suficiente carbonato de calcio disponible para construir un caparazón realmente sólido, como en algunos suelos ácidos de la tierra, aparecen varias especies de babosas y también algunos caracoles con caparazones delgados y translúcidos, compuestos mayoritariamente o totalmente por la proteína conquiolina .
Los caracoles como Sphincterochila boissieri y Xerocrassa seetzeni se han adaptado a las condiciones del desierto. Otros caracoles se han adaptado a vivir en zanjas, cerca de fuentes hidrotermales de aguas profundas , en fosas oceánicas a 10.000 metros (6 millas) por debajo de la superficie, [17] en las fuertes olas de las costas rocosas , cuevas y muchas otras áreas diversas.
Los gasterópodos pueden ser transferidos accidentalmente de un hábitat a otro por otros animales, por ejemplo, las aves . [18]
Anatomía
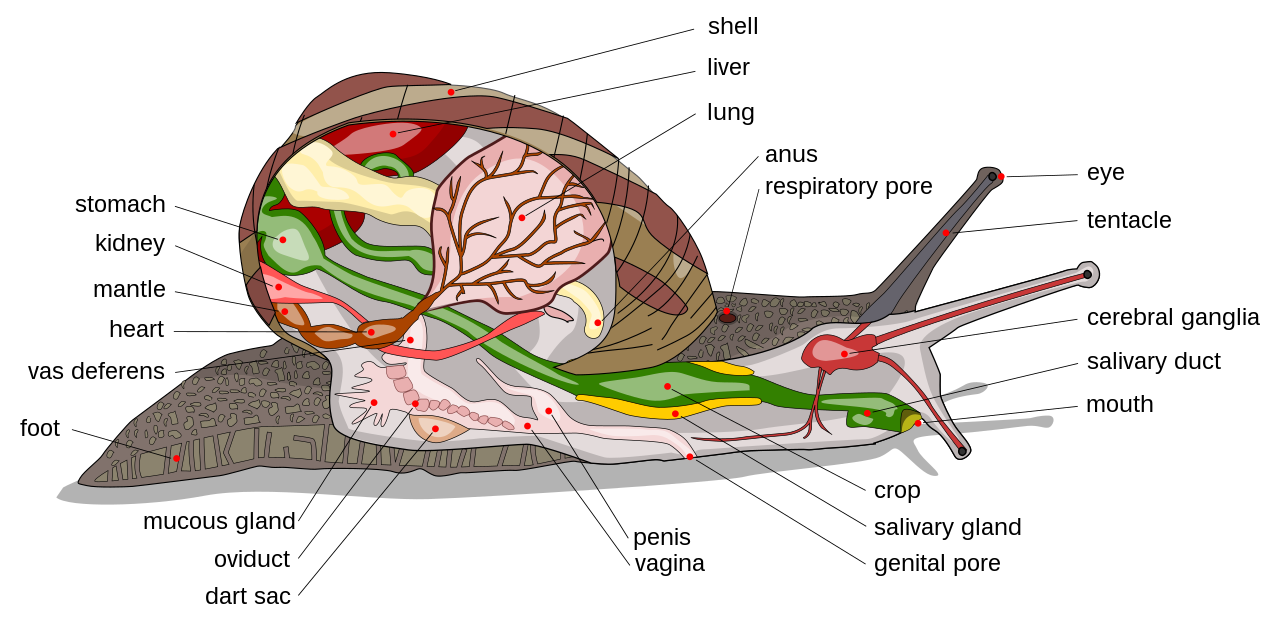
Los caracoles se distinguen por un proceso anatómico conocido como torsión , en el que la masa visceral del animal gira 180° hacia un lado durante el desarrollo, de modo que el ano se sitúa más o menos por encima de la cabeza. Este proceso no está relacionado con el enrollamiento de la concha, que es un fenómeno independiente. La torsión está presente en todos los gasterópodos, pero los gasterópodos opistobranquios no están torcidos en diversos grados. [19] [20]
La torsión se produce en dos etapas. La primera, mecanicista, es muscular y la segunda es mutagénica. Los efectos de la torsión son principalmente fisiológicos. El organismo se desarrolla mediante un crecimiento asimétrico, y la mayor parte del crecimiento se produce en el lado izquierdo. Esto conduce a la pérdida de la anatomía del lado derecho, que en la mayoría de los bilaterales es un duplicado de la anatomía del lado izquierdo. La característica esencial de esta asimetría es que el ano generalmente se encuentra a un lado del plano medio. Las crestas branquiales , los órganos olfativos , la glándula mucosa del pie , los nefridios y la aurícula del corazón son únicos o al menos están más desarrollados en un lado del cuerpo que en el otro. Además, solo hay un orificio genital , que se encuentra en el mismo lado del cuerpo que el ano. [21] Además, el ano se redirige al mismo espacio que la cabeza. Se especula que esto tiene alguna función evolutiva, ya que antes de la torsión, al retraerse dentro del caparazón, primero se retraía el extremo posterior y luego el anterior. Ahora, el extremo frontal se puede retraer con mayor facilidad, lo que tal vez sugiera un propósito defensivo.
Los gasterópodos suelen tener una cabeza bien definida con dos o cuatro tentáculos sensoriales con ojos y un pie ventral. La primera división del pie se llama propodio. Su función es empujar los sedimentos mientras el caracol se arrastra. La concha larval de un gasterópodo se llama protoconcha .
Caparazón
.jpg/1280px-Zonitoides_nitidus_shell_(cropped).jpg)
La mayoría de los gasterópodos con concha tienen una concha de una sola pieza (con excepción de los gasterópodos bivalvos ), típicamente enrollada o en espiral, al menos en la etapa larvaria. Esta concha enrollada generalmente se abre en el lado derecho (visto con el ápice de la concha apuntando hacia arriba). Numerosas especies tienen un opérculo , que en muchas especies actúa como una trampilla para cerrar la concha. Este suele estar hecho de un material similar al cuerno, pero en algunos moluscos es calcáreo. En las babosas terrestres, la concha es reducida o ausente, y el cuerpo es aerodinámico.
Algunos gasterópodos tienen conchas adultas que son pesadas en la parte inferior debido a la presencia de un depósito de callo ventral grueso, a menudo ancho y convexo en el labio interno y adápico a la abertura, lo que puede ser importante para la estabilidad gravitacional. [22]
Pared del cuerpo
Algunas babosas marinas tienen colores muy llamativos. Esto sirve como advertencia , cuando son venenosas o contienen células urticantes , o para camuflarlas en los hidroides , esponjas y algas de colores brillantes en los que se encuentran muchas de las especies.
Las excrecencias laterales del cuerpo de los nudibranquios se denominan cerata y contienen unas evaginaciones de glándulas digestivas llamadas divertículos .
Órganos sensoriales y sistema nervioso

Los órganos sensoriales de los gasterópodos incluyen órganos olfativos , ojos, estatocistos y mecanorreceptores . [23] Los gasterópodos no tienen audición. [23]
En los gasterópodos terrestres (caracoles terrestres y babosas), los órganos olfativos, ubicados en las puntas de los cuatro tentáculos , son el órgano sensorial más importante. [23] Los órganos quimiosensoriales de los gasterópodos marinos opistobranquios se denominan rinóforos .
La mayoría de los gasterópodos tienen órganos visuales simples, manchas oculares ya sea en la punta o en la base de los tentáculos. Sin embargo, los "ojos" en los gasterópodos varían desde simples ocelos que solo distinguen claro y oscuro, hasta ojos de fosetas más complejos , e incluso ojos de cristalino . [24] En los caracoles terrestres y las babosas, la visión no es el sentido más importante, porque son animales principalmente nocturnos . [23]
El sistema nervioso de los gasterópodos incluye el sistema nervioso periférico y el sistema nervioso central . El sistema nervioso central está formado por ganglios conectados por células nerviosas. Incluye ganglios pares: ganglios cerebrales, ganglios pedios, ganglios osphradiales , ganglios pleurales, ganglios parietales y ganglios viscerales. A veces también hay ganglios bucales. [23]
Sistema digestivo
La rádula de un gasterópodo suele estar adaptada al alimento que come una especie. Los gasterópodos más simples son las lapas y las abulones , herbívoros que utilizan su rádula dura para raspar las algas de las rocas.
Muchos gasterópodos marinos son excavadores y tienen un sifón que se extiende desde el borde del manto . A veces, la concha tiene un canal sifonal para acomodar esta estructura. Un sifón permite al animal atraer agua hacia la cavidad del manto y por las branquias. Utilizan el sifón principalmente para "probar" el agua y detectar presas a distancia. Los gasterópodos con sifón tienden a ser depredadores o carroñeros.
Sistema respiratorio
Casi todos los gasterópodos marinos respiran con branquias , pero muchas especies de agua dulce y la mayoría de las especies terrestres tienen un pulmón paleal . La proteína respiratoria en casi todos los gasterópodos es la hemocianina , pero una familia de pulmonados de agua dulce, los Planorbidae , tiene hemoglobina como proteína respiratoria.
En un gran grupo de babosas marinas, las branquias están dispuestas como una roseta de penachos plumosos en el dorso, lo que da lugar a su otro nombre, nudibranquios . Algunos nudibranquios tienen el dorso liso o verrugoso sin mecanismo branquial visible, de modo que es probable que la respiración se realice directamente a través de la piel.
Sistema circulatorio
Los gasterópodos tienen un sistema circulatorio abierto y el fluido de transporte es la hemolinfa . La hemocianina está presente en la hemolinfa como pigmento respiratorio.
Sistema excretor
Los principales órganos de excreción de los gasterópodos son los nefridios , que producen amoníaco o ácido úrico como producto de desecho. El nefridio también desempeña un papel importante en el mantenimiento del equilibrio hídrico en especies terrestres y de agua dulce. Otros órganos de excreción, al menos en algunas especies, incluyen las glándulas pericárdicas en la cavidad corporal y las glándulas digestivas que desembocan en el estómago .
Sistema reproductivo

El cortejo es parte del comportamiento de apareamiento en algunos gasterópodos, incluidos algunos de los helícidos . Además, en algunos caracoles terrestres, una característica inusual del sistema reproductivo de los gasterópodos es la presencia y utilización de dardos amorosos .
En muchos gasterópodos marinos distintos de los opistobranquios , existen sexos separados ( dioicos / gonocóricos ); la mayoría de los gasterópodos terrestres, sin embargo, son hermafroditas .
Ciclo vital

El cortejo es parte del comportamiento de apareamiento de los gasterópodos con algunas familias pulmonadas de caracoles terrestres que crean y utilizan dardos de amor , cuyo lanzamiento se ha identificado como una forma de selección sexual . [25]
Los principales aspectos del ciclo de vida de los gasterópodos incluyen:
- La puesta de huevos y los huevos de los gasterópodos
- El desarrollo embrionario de los gasterópodos
- Las larvas o estadio larvario: algunos gasterópodos pueden ser trocóforos y/o veligeros.
- Estivación e hibernación (cada una de estas está presente sólo en algunos gasterópodos)
- El crecimiento de los gasterópodos
- Cortejo y apareamiento en gasterópodos: la fecundación es interna o externa según la especie. La fecundación externa es común en los gasterópodos marinos.
Comportamiento alimentario

La dieta de los gasterópodos difiere según el grupo considerado. Los gasterópodos marinos incluyen algunos que son herbívoros , detritívoros , carnívoros depredadores , carroñeros , parásitos y también unos pocos ciliares, en los que la rádula está reducida o ausente. Las especies terrestres pueden masticar hojas, corteza, fruta y animales en descomposición, mientras que las especies marinas pueden raspar algas de las rocas del fondo marino. Ciertas especies, como Archaeogastropda, mantienen filas horizontales de dientes marginales delgados. En algunas especies que han evolucionado hasta convertirse en endoparásitos, como el eulimido Thyonicola doglieli , muchas de las características estándar de los gasterópodos están muy reducidas o ausentes.
Algunas babosas marinas son herbívoras y otras son carnívoras. El hábito carnívoro se debe a la especialización. Muchos gasterópodos tienen preferencias alimentarias específicas y suelen estar en estrecha asociación con las especies de las que se alimentan.
Algunos gasterópodos carnívoros depredadores incluyen: conchas cónicas , Testacella , Daudebardia , túrridos , babosas fantasmas y otros.
Genética
Los gasterópodos exhiben un grado importante de variación en la organización de genes mitocondriales en comparación con otros animales. [26] Los principales eventos de reordenamiento genético ocurrieron en el origen de Patellogastropoda y Heterobranchia , mientras que ocurrieron menos cambios entre los ancestros de Vetigastropoda (solo ARNt D, C y N) y Caenogastropoda (una gran inversión única y translocaciones de los ARNt D y N). [26] Dentro de Heterobranchia, el orden genético parece relativamente conservado, y los reordenamientos genéticos están relacionados principalmente con la transposición de genes de ARNt. [26]
Historia y evolución geológica


Los primeros gasterópodos fueron exclusivamente marinos, y los primeros representantes del grupo aparecieron en el Cámbrico tardío ( Chippewaella , Strepsodiscus ), [27] aunque su único carácter gasterópodo es una concha enrollada, por lo que podrían estar en el linaje del tallo, si es que son gasterópodos. [28] Los primeros organismos cámbricos como Helcionella , Barskovia y Scenella ya no se consideran gasterópodos, [ cita requerida ] y la diminuta Aldanella enrollada del Cámbrico más temprano probablemente ni siquiera sea un molusco. [ cita requerida ]
Por lo tanto, no es hasta el Ordovícico que surgen los primeros miembros del grupo corona. [29] En el período Ordovícico, los gasterópodos eran un grupo variado presente en una variedad de hábitats acuáticos. Comúnmente, los gasterópodos fósiles de las rocas de la era Paleozoica temprana están demasiado mal conservados para una identificación precisa. Aún así, el género silúrico Poleumita contiene quince especies identificadas. Los gasterópodos fósiles fueron menos comunes durante la era Paleozoica que los bivalvos . [29]
La mayoría de los gasterópodos de la era Paleozoica pertenecen a grupos primitivos, de los cuales algunos aún sobreviven. En el período Carbonífero , muchas de las formas que se observan en los gasterópodos actuales pueden coincidir con el registro fósil, pero a pesar de estas similitudes en apariencia, la mayoría de estas formas más antiguas no están directamente relacionadas con las formas actuales. Fue durante la era Mesozoica cuando evolucionaron los ancestros de muchos de los gasterópodos actuales. [29]
Uno de los primeros gasterópodos terrestres (que habitaban en la tierra) conocidos es Anthracopupa (= Maturipupa ), [30] que se encuentra en las cuencas carboníferas del período Carbonífero en Europa, pero los parientes de los caracoles terrestres modernos son raros antes del período Cretácico . [29]
En las rocas de la era Mesozoica , los gasterópodos son ligeramente más comunes como fósiles; sus conchas suelen estar bien conservadas. Sus fósiles aparecen en antiguos lechos depositados tanto en ambientes de agua dulce como marinos. El « mármol de Purbeck » del período Jurásico y el « mármol de Sussex » del período Cretácico temprano, que se encuentran ambos en el sur de Inglaterra, son calizas que contienen los restos compactados del caracol de estanque Viviparus . [29]
Las rocas de la era Cenozoica contienen una gran cantidad de fósiles de gasterópodos, muchos de los cuales están estrechamente relacionados con las formas de vida modernas. La diversidad de los gasterópodos aumentó notablemente al comienzo de esta era, junto con la de los bivalvos . [29]
Se cree que ciertas marcas similares a rastros preservadas en rocas sedimentarias antiguas fueron hechas por gasterópodos que se arrastraban sobre el barro blando y la arena. Aunque estos rastros fósiles son de origen discutible, algunos de ellos se parecen a los rastros hechos por los gasterópodos actuales. [29]
Los fósiles de gasterópodos a veces pueden confundirse con amonitas u otros cefalópodos con concha . Un ejemplo de esto es Belerofonte de las calizas del período Carbonífero en Europa, cuya concha está enrollada en espiral y puede confundirse con la concha de un cefalópodo. [ cita requerida ]
Los gasterópodos son uno de los grupos que registran los cambios en la fauna provocados por el avance y retroceso de las capas de hielo durante el Pleistoceno .
Cladograma
Un cladograma que muestra las relaciones filogenéticas de Gastropoda con especies de ejemplo: [31]
| Gasterópodos | |

.jpg/1280px-Ancylus_fluviatilis_O.F.M%C3%BCller,_1774_(4206383684).jpg)




_(2999358117).jpg/1280px-Ampullaria_ampullacea_(Linn%C3%A9)_(2999358117).jpg)
_(4309322782).jpg/1280px-Acicula_lineata_(Draparnaud,_1801)_(4309322782).jpg)


_(3063441884).jpg/1280px-Acanthina_brevidentata_(Wood,_1828)_(3063441884).jpg)


_(3000195752).jpg/1280px-Theodoxus_anatolicus_(R%C3%A9cluz,_1841)_(3000195752).jpg)


_(4947706383).jpg/1280px-Lucapina_suffusa_(Reeve,_1850)_(4947706383).jpg)
.jpg/1280px-Fissurella_nimbosa_Linn%C3%A9,_1758_(4426866577).jpg)
_(4631972937).jpg/1280px-Patelloida_pustulata_(Helbling,_1779)_(4631972937).jpg)
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
_(2999358117).jpg){kind=link}
_(4309322782).jpg){kind=link}
{kind=link}
{kind=link}
_(3063441884).jpg){kind=link}
{kind=link}
{kind=link}
_(3000195752).jpg){kind=link}
{kind=link}
{kind=link}
_(4947706383).jpg){kind=link}
.jpg){kind=link}
_(4631972937).jpg){kind=link}
Cocculiniformia , Neomphalina y Lower Heterobranchia no están incluidos en el cladograma anterior.
Taxonomía
Clasificación actual
La clasificación actual de los gasterópodos se basa en los resultados de los análisis filogenómicos. No se ha llegado a un consenso aún considerando las relaciones en la base del árbol de la vida de los gasterópodos, pero por lo demás se conocen con seguridad los grupos principales. [32] [33] [34]
- Gasterópodos
- Adenogonogastropoda [33] (Angiogastropoda [32] )
- Patelogastrópodos
- Vetigastropoda (incluidos Neomphaliones )
{kind=link}

{kind=link}

{kind=link}

Historia
Desde Darwin , la taxonomía biológica ha intentado reflejar la filogenia de los organismos, es decir, el árbol de la vida . Las clasificaciones utilizadas en taxonomía intentan representar la interrelación precisa de los diversos taxones. Sin embargo, la taxonomía de los gasterópodos se revisa constantemente y, por lo tanto, las versiones que se muestran en los distintos textos pueden diferir en aspectos importantes.
En la clasificación más antigua de los gasterópodos, había cuatro subclases: [35]
- Opistobranquios (branquias a la derecha y detrás del corazón).
- Gymnomorpha (sin caparazón)
- Prosobranquias (branquias delante del corazón).
- Pulmonata (con pulmón en lugar de branquias)
La taxonomía de los gasterópodos todavía se encuentra en revisión y cada vez se abandonan más conceptos antiguos a medida que los resultados de los estudios de ADN se vuelven más claros. No obstante, algunos términos antiguos, como "opistobranquio" y "prosobranquio", todavía se utilizan a veces de manera descriptiva.
Los nuevos conocimientos basados en la secuenciación del ADN de los gasterópodos han producido algunos nuevos y revolucionarios conocimientos taxonómicos. En el caso de los gasterópodos, la taxonomía se está reescribiendo gradualmente para incorporar grupos estrictamente monofiléticos (solo un linaje de gasterópodos en cada grupo). La integración de los nuevos hallazgos en una taxonomía funcional sigue siendo un desafío. Los rangos consistentes dentro de la taxonomía a nivel de subclase, superorden, orden y suborden ya se han abandonado por considerarlos inviables. Se esperan revisiones continuas de los niveles taxonómicos superiores en el futuro cercano. [ ¿Cuándo? ]
La evolución convergente , que parece existir con una frecuencia especialmente alta en los gasterópodos, puede explicar las diferencias observadas entre las filogenias más antiguas, que se basaban en datos morfológicos, y los estudios de secuenciación genética más recientes.
En 2004, Brian Simison y David R. Lindberg demostraron posibles orígenes difiléticos de los Gastropoda basándose en el orden de los genes mitocondriales y en análisis de la secuencia de aminoácidos de genes completos. [36]
En 2005, Philippe Bouchet y Jean-Pierre Rocroi realizaron cambios radicales en la sistemática , lo que resultó en la taxonomía Bouchet & Rocroi , que es un paso más hacia la historia evolutiva del filo . [3] [37] El sistema de clasificación Bouchet & Rocroi se basa en parte en los sistemas de clasificación más antiguos y en parte en la nueva investigación cladística . En el pasado, la taxonomía de los gasterópodos se basaba en gran medida en caracteres morfológicos fenéticos de los taxones. Los avances recientes se basan más en caracteres moleculares del ADN [38] y la investigación del ARN . Esto ha hecho que los rangos taxonómicos y su jerarquía sean controvertidos.
En 2017, Bouchet, Rocroi y otros colaboradores publicaron una versión significativamente actualizada de la taxonomía de 2005. [39] En la taxonomía de Bouchet et al. , los autores utilizaron clados no clasificados para taxones por encima del rango de superfamilia (reemplazando los rangos suborden, orden, superorden y subclase), mientras que utilizaron el enfoque tradicional de Linneo para todos los taxones por debajo del rango de superfamilia. Siempre que no se ha probado la monofilia , o se sabe que es parafilética o polifilética, se ha utilizado el término "grupo" o "grupo informal". La clasificación de familias en subfamilias a menudo no está bien resuelta. [ cita requerida ]
Conservación
Muchas especies de gasterópodos se enfrentan a amenazas derivadas de la destrucción del hábitat, la contaminación y el cambio climático. Algunas especies están en peligro o se han extinguido debido a estos factores. Los esfuerzos de conservación suelen centrarse en proteger sus hábitats, especialmente en los ecosistemas terrestres y de agua dulce.
Referencias
- ^ Aktipis, SO; Giribet, G.; Lindberg, DR; WF, reflexionar (2008). "Gastropoda". En Ponder, W.; Lindberg, DR (eds.). Filogenia y evolución de los Moluscos . Prensa de la Universidad de California. págs. 201–238. ISBN 978-0-520-25092-5.
- ^ ab Cuvier, G (1795). "Second mémoire sur l'organisation et les rapports des animaux à sang blanc, dans lequel on tracee de la Structure des Mollusques et de leur division en ordres, lu à la Société d'histoire Naturelle de Paris, le 11 Prairial, an III" . Magazin Encyclopédique, ou Journal des Sciences, des Lettres et des Arts (en francés). 2 : 433–449. Archivado desde el original el 25 de julio de 2017.
- ^ abc Bouchet P. & Rocroi J.-P. (Ed.); Frýda J., Hausdorf B., Ponder W., Valdes A. & Warén A. 2005. Clasificación y nomenclador de familias de gasterópodos . Malacologia: Revista internacional de malacología, 47(1-2). ConchBooks: Hackenheim, Alemania. ISBN 3-925919-72-4 . 397 pp. vliz.be Archivado el 14 de julio de 2020 en Wayback Machine.
- ^ ab Solem, AG "Gastropod". Encyclopaedia Britannica . Encyclopaedia Britannica Inc. Archivado desde el original el 8 de marzo de 2016 . Consultado el 6 de marzo de 2017 .
- ^ ab Bieler R, Bouchet P, Gofas S, Marshall B, Rosenberg G, La Perna R, Neubauer TA, Sartori AF, Schneider S, Vos C, ter Poorten JJ, Taylor J, Dijkstra H, Finn J, Bank R, Neubert E, Moretzsohn F, Faber M, Houart R, Picton B, García-Álvarez O, eds. (2020). "Gastropoda". Base de moluscos . Registro Mundial de Especies Marinas . Consultado el 29 de septiembre de 2020 .
- ^ Strong, Ellen E.; Schrödl, Michael; Parkhaev, Pavel; Nützel, Alexander; Kano, Yasunori; Kaim, Andrzej; Hausdorf, Bernhard; Rocroi, Jean-Pierre; Bouchet, Philippe (31 de diciembre de 2017). "Clasificación revisada, nomenclador y tipificación de familias de gasterópodos y monoplacóforos". Malacología . 61 (1–2): 1–526. doi :10.4002/040.061.0201. S2CID 91051256.
- ^ "Gasterópodo". Diccionario Etimológico en Línea . Archivado desde el original el 6 de octubre de 2018. Consultado el 3 de septiembre de 2019 .
- ^ McArthur, AG; MG Harasewych (2003). "Sistemática molecular de los principales linajes de los gasterópodos". En C. Lydeard; DR Lindberg (eds.). Sistemática molecular y filogeografía de los moluscos . Washington: Smithsonian Books. págs. 140–160.
- ^ ab Chapman, AD (2009). Número de especies vivas en Australia y el mundo, 2.ª edición Archivado el 6 de diciembre de 2010 en Wayback Machine . Estudio de recursos biológicos de Australia, Canberra. Consultado el 12 de enero de 2010. ISBN 978-0-642-56860-1 (impreso); ISBN 978-0-642-56861-8 (en línea).
- ^ Appeltans W., Bouchet P., Boxshall GA, Fauchald K., Gordon DP, Hoeksema BW, Poore GCB, van Soest RWM, Stöhr S., Walter TC, Costello MJ (eds) (2011). Registro Mundial de Especies Marinas. Consultado en marinespecies.org. Archivado el 3 de octubre de 2015 en archive.today el 7 de marzo de 2011.
- ^ "Censo de la vida marina (2012). SYNDEEP: Hacia una primera síntesis global de biodiversidad, biogeografía y función ecosistémica en las profundidades marinas. Datos no publicados (datasetID: 59)" (PDF) . Archivado (PDF) desde el original el 2019-06-30 . Consultado el 2019-04-25 .
- ^ "gasterópodo" Archivado el 13 de octubre de 2008 en Wayback Machine . (2010). En Encyclopædia Britannica. Recuperado el 5 de marzo de 2010 de Encyclopædia Britannica Online.
- ^ Strong, Ellen E.; Gargominy, Olivier; Ponder, Winston F.; Bouchet, Philippe (enero de 2008). "Diversidad global de gasterópodos (Gastropoda; Mollusca) en agua dulce". Hydrobiologia . 595 (1): 149–166. Bibcode :2008HyBio.595..149S. doi :10.1007/s10750-007-9012-6. hdl :10088/7390. S2CID 44234861.
- ^ Régnier, C.; Fontaine, BT; Bouchet, P. (octubre de 2009). "No saber, no registrar, no enumerar: numerosas extinciones de moluscos inadvertidas". Biología de la conservación . 23 (5): 1214–1221. Bibcode :2009ConBi..23.1214R. doi :10.1111/j.1523-1739.2009.01245.x. PMID 19459894. S2CID 8021609.
- ^ (en español) Nájera JM (1996). "Moluscos del suelo como plagas agrícolas y cuarentenarias". X Congreso Nacional Agronómico / II Congreso de Suelos 1996 51-56. PDF Archivado el 21 de julio de 2011 en Wayback Machine.
- ^ Rex, Michael A. (14 de septiembre de 1973). "Diversidad de especies de aguas profundas: disminución de la diversidad de gasterópodos en profundidades abisales". Science . 181 (4104): 1051–1053. Bibcode :1973Sci...181.1051R. doi :10.1126/science.181.4104.1051. ISSN 0036-8075. PMID 17731267. S2CID 43572097.
- ^ Fukumori, Hiroaki; Takano, Tsuyoshi; Hasegawa, Kazunori; Kano, Yasunori (2019). "La fauna de gasterópodos más profunda conocida: composición y distribución de especies en la fosa de Kuril-Kamchatka". Progress in Oceanography . 178 : 102176. Bibcode :2019PrOce.17802176F. doi :10.1016/j.pocean.2019.102176.
- ^ RUSIECKI S. & RUSIECKA A. 2013. Caracol peludo Trochulus hispidus (Linnaeus, 1758) en vuelo: una nota sobre la dispersión de caracoles por las aves. Archivado el 16 de septiembre de 2013 en Wayback Machine. Folia Malacologica 21(2):111-112.
- ^ Kay, A.; Wells, FE; Poder, WF (1998). "Clase Gastropoda". En Beesley, PL; Ross, GJB; Wells, A. (eds.). Mollusca: The Southern Synthesis. Fauna de Australia . CSIRO Publishing. págs. 565–604. ISBN 978-0-643-05756-2.
- ^ Brusca, RC; Brusca, GJ (2003). "Filo Mollusca". Invertebrados . Sinauer Associates, Inc., págs. 701–769. ISBN 978-0-87893-097-5.
- ^ Suter, Henry. Manual de los moluscos de Nueva Zelanda /. J. Mackay, impresor del gobierno. Archivado desde el original el 26 de septiembre de 2017. Consultado el 14 de enero de 2018 .
- ^ Geerat J. Vermeij (2022). «La vida equilibrada: evolución del peso de la concha ventral en los gasterópodos». Revista Zoológica de la Sociedad Linneana . 194 (1): 256–275. doi :10.1093/zoolinnean/zlab019. Archivado desde el original el 23 de agosto de 2023. Consultado el 4 de enero de 2022 .
- ^ abcde Chase R.: Órganos sensoriales y sistema nervioso . en Barker GM (ed.): La biología de los moluscos terrestres . CABI Publishing, Oxon, Reino Unido, 2001, ISBN 0-85199-318-4 . 1-146, páginas citadas: 179–211.
- ^ Götting, Klaus-Jürgen (1994). "Schnecken". En Becker, U.; Ganter, S.; Justo, C.; Sauermost, R. (eds.). Lexikon der Biologie . Heidelberg: Spektrum Akademischer Verlag. ISBN 978-3-86025-156-0.
- ^ Koene, Joris M. (1 de agosto de 2006). «Cuentos de dos caracoles: selección sexual y conflicto sexual en Lymnaea stagnalis y Helix aspersa». Biología Integrativa y Comparada . 46 (4): 419–429. doi : 10.1093/icb/icj040 . PMID 21672754. Archivado desde el original el 1 de septiembre de 2019. Consultado el 31 de agosto de 2019 en academic.oup.com.
- ^ abc Cunha, RL; Grande, C.; Zardoya, R. (2009). "Relaciones filogenéticas de neogastropodos basadas en genomas mitocondriales completos". BMC Evolutionary Biology . 9 (1): 210. Bibcode :2009BMCEE...9..210C. doi : 10.1186/1471-2148-9-210 . PMC 2741453 . PMID 19698157.
- ^ Yochelson, Ellis L.; Nuelle, Laurence M. (1985). "Strepsodiscus (Gastropoda) en el Cámbrico Tardío de Missouri". Revista de Paleontología . 59 (3): 733–740. JSTOR 1304993.
- ^ Budd, GE y S. Jensen. 2000: Una reevaluación crítica del registro fósil de los filos bilaterales. Biological Reviews 75:253–295.
- ^ abcdefg Be'Norr, K., y J. FnÍon (1999). «Notas sobre la evolución y clasificación superior de la subclase Neritimorpha (Gastropoda) con la descripción de algunos taxones nuevos» (PDF) . Geol. Et Palaeont 33 : 219–235. Archivado (PDF) desde el original el 13 de diciembre de 2017. Consultado el 13 de diciembre de 2017 .
{{cite journal}}: CS1 maint: varios nombres: lista de autores ( enlace ) - ^ Jochum, Adrienne; Yu, Tingting; Neubauer, Thomas A. (2020). "Primer registro de la familia de caracoles terrestres del Paleozoico Anthracopupidae en el Jurásico Inferior de China y el origen de Stylommatophora". Revista de Paleontología . 94 (2): 266–272. Código Bibliográfico :2020JPal...94..266J. doi :10.1017/jpa.2019.68. ISSN 0022-3360. S2CID 204269735. Archivado desde el original el 23 de agosto de 2023 . Consultado el 29 de julio de 2022 .
- ^ Kenny, Nathan J.; Truchado-García, Marta; Grande, Cristina (2016). "El transcriptoma profundo y multietapa del vector de la esquistosomiasis Biomphalaria glabrata proporciona una plataforma para comprender las vías relacionadas con la enfermedad de los moluscos". BMC Infectious Diseases . 16 (1): 618. doi : 10.1186/s12879-016-1944-x . ISSN 1471-2334. PMC 5084317 . PMID 27793108.
- ^ ab Cunha, Tauana Junqueira; Giribet, Gonzalo (13 de marzo de 2019). "Una topología congruente para las relaciones profundas entre gasterópodos". Actas de la Royal Society B: Ciencias Biológicas . 286 (1898): 20182776. doi :10.1098/rspb.2018.2776. ISSN 0962-8452. PMC 6458328 . PMID 30862305.
- ^ ab Uribe, Juan E; González, Vanessa L; Irisarri, Iker; Kano, Yasunori; Herbert, David G; Strong, Ellen E; Harasewych, MG (12 de octubre de 2022). Oaks, Jamie (ed.). "Una columna vertebral filogenómica para los moluscos gasterópodos". Biología sistemática . 71 (6): 1271–1280. doi :10.1093/sysbio/syac045. hdl : 10261/279410 . ISSN 1063-5157. PMID 35766870. Archivado desde el original el 23 de agosto de 2023 . Consultado el 10 de agosto de 2023 .
- ^ Chen, Zeyuan; Schrödl, Michael (18 de abril de 2022). "¿Cuántos genes ortólogos de una sola copia de genomas completos revelan relaciones profundas entre gasterópodos?". PeerJ . 10 : e13285. doi : 10.7717/peerj.13285 . ISSN 2167-8359. PMC 9048639 . PMID 35497189.
- ^ Paul Jeffery. Clasificación supragenérica de la clase Gastropoda . Museo de Historia Natural, Londres, 2001.
- ^ "Unitas malacologica, Boletín número 21 de diciembre de 2004 - archivo .pdf" (PDF) . Archivado (PDF) desde el original el 15 de junio de 2011 . Consultado el 9 de marzo de 2009 .
- ^ Poppe GT & Tagaro SP 2006. La nueva clasificación de los gasterópodos según Bouchet & Rocroi, 2005. Visaya, febrero de 2006: 10 pp. journal-malaco.fr Archivado el 27 de septiembre de 2007 en Wayback Machine.
- ^ Elpidio A. Remigio y Paul DN Hebert (2003). "Prueba de la utilidad de secuencias parciales de COI para la filogenia (texto completo en línea)" (PDF) . Filogenética molecular y evolución . 29 (3): 641–647. doi :10.1016/S1055-7903(03)00140-4. PMID 14615199. Archivado desde el original (PDF) el 27 de septiembre de 2007.
- ^ Philippe Bouchet, Jean-Pierre Rocroi, Bernhard Hausdorf, Andrzej Kaim, Yasunori Kano, Alexander Nützel, Pavel Parkhaev, Michael Schrödl y Ellen E. Strong. 2017. Clasificación revisada, nomenclador y tipificación de familias de gasterópodos y monoplacóforos Archivado el 23 de agosto de 2023 en Wayback Machine . Malacología , 61(1-2): 1-526.
Fuentes
- Este artículo incorpora texto CC-BY-2.0 de la siguiente fuente: Cunha, RL; Grande, C.; Zardoya, R. (2009). "Relaciones filogenéticas de neogastropodos basadas en genomas mitocondriales completos". BMC Evolutionary Biology . 9 : 210. doi :10.1186/1471-2148-9-210. PMC 2741453. PMID 19698157.
- Abbott, RT (1989): Compendio de conchas terrestres. Una guía en color de más de 2000 de las conchas terrestres del mundo. 240 S., American Malacologists. Melbourne, Fl, Burlington, Ma. ISBN 0-915826-23-2
- Abbott, RT y Dance, SP (1998): Compendio de conchas marinas. Una guía a todo color de más de 4200 conchas marinas del mundo. 413 S., Odyssey Publishing. El Cajon, California. ISBN 0-9661720-0-0
- Parkinson, B., Hemmen, J. y Groh, K. (1987): Conchas terrestres tropicales del mundo. 279 S., editorial Christa Hemmen. Wiesbaden. ISBN 3-925919-00-7
- Ponder, WF y Lindberg, DR (1997): Hacia una filogenia de los moluscos gasterópodos: un análisis utilizando caracteres morfológicos . Zoological Journal of the Linnean Society , 119 83–265.
- Robin, A. (2008): Enciclopedia de gasterópodos marinos. 480 págs., Editorial ConchBooks. Hackenheim. ISBN 978-3-939767-09-1
Enlaces externos
- Comportamiento reproductivo de los gasterópodos
- Taxonomía linneana de gasterópodos de 2004
- Webster, S.; Fiorito, G. (2001). "Comportamiento socialmente guiado en invertebrados no insectos". Animal Cognition . 4 (2): 69. doi :10.1007/s100710100108. S2CID 25373798.– Artículo sobre el aprendizaje social también en gasterópodos.
- Galería de fotografías de gasterópodos, principalmente fósiles, algunas conchas modernas
- Un vídeo de un caracol de jardín arrastrándose ( Cornu aspersum ), YouTube
- Grove, SJ (2018). Una guía sobre las conchas y otros moluscos marinos de Tasmania: Moluscos de Tasmania con imágenes
