Opsina visual de vertebrados
This article needs additional citations for verification. (November 2022) |

{kind=link}
{kind=link}

Las opsinas visuales de vertebrados son una subclase de las opsinas ciliares y median la visión en los vertebrados. Incluyen las opsinas de los bastones y conos humanos . A menudo se las abrevia como opsinas , ya que fueron las primeras opsinas descubiertas y siguen siendo las opsinas más estudiadas. [1]
Opsinas
La opsina se refiere estrictamente a la apoproteína (sin retinal unido). Cuando una opsina se une al retinal para formar una holoproteína , se la denomina proteína retinilideno . Sin embargo, a menudo se ignora la distinción y la opsina puede referirse vagamente a ambas (independientemente de si el retinal está unido o no).
Las opsinas son receptores acoplados a proteína G (GPCR) y deben unirse al retinal —normalmente 11- cis -retinal — para ser fotosensibles, ya que el retinal actúa como cromóforo . Cuando la proteína retinilideno absorbe un fotón, el retinal se isomeriza y es liberado por la opsina. El proceso que sigue a la isomerización y renovación del retinal se conoce como ciclo visual . El 11- cis -retinal libre es fotosensible y tiene su propia sensibilidad espectral de 380 nm. [2] Sin embargo, para desencadenar la cascada de fototransducción , el proceso que subyace a la señal visual, el retinal debe estar unido a una opsina cuando se isomeriza. La proteína retinilideno tiene una sensibilidad espectral que difiere de la del retinal libre y depende de la secuencia de opsina.
Si bien las opsinas solo pueden unirse al retinal, existen dos formas de retinal que pueden actuar como cromóforo para las opsinas visuales de los vertebrados:
- Retinal 1 ( 11- cis -Retinal ): la forma común presente en la mayoría de las opsinas
- Retinal 2 ( 11- cis -3,4-dehidroretinal ): una forma más rara que está relativamente desplazada hacia el rojo en comparación con el retinal 1.
Los animales que viven en tierra y los peces marinos forman sus pigmentos visuales exclusivamente con retinal 1. Sin embargo, muchos peces de agua dulce y anfibios también pueden formar pigmentos visuales con retinal 2, dependiendo de la activación de la enzima retinal-3,4-desaturasa (GO:0061899). Muchas de estas especies pueden cambiar entre estos cromóforos durante su ciclo de vida, para adaptarse a un hábitat cambiante. [3] [4]
Función
{kind=link}
La isomerización del 11- cis -retinal en todo- trans -retinal por la luz induce un cambio conformacional en la proteína que activa la vía de fototransducción .
Subclases
Hay dos clases de opsina visual en vertebrados, que se diferencian según se expresen en fotorreceptores de bastones o de conos.
Opsinas cónicas
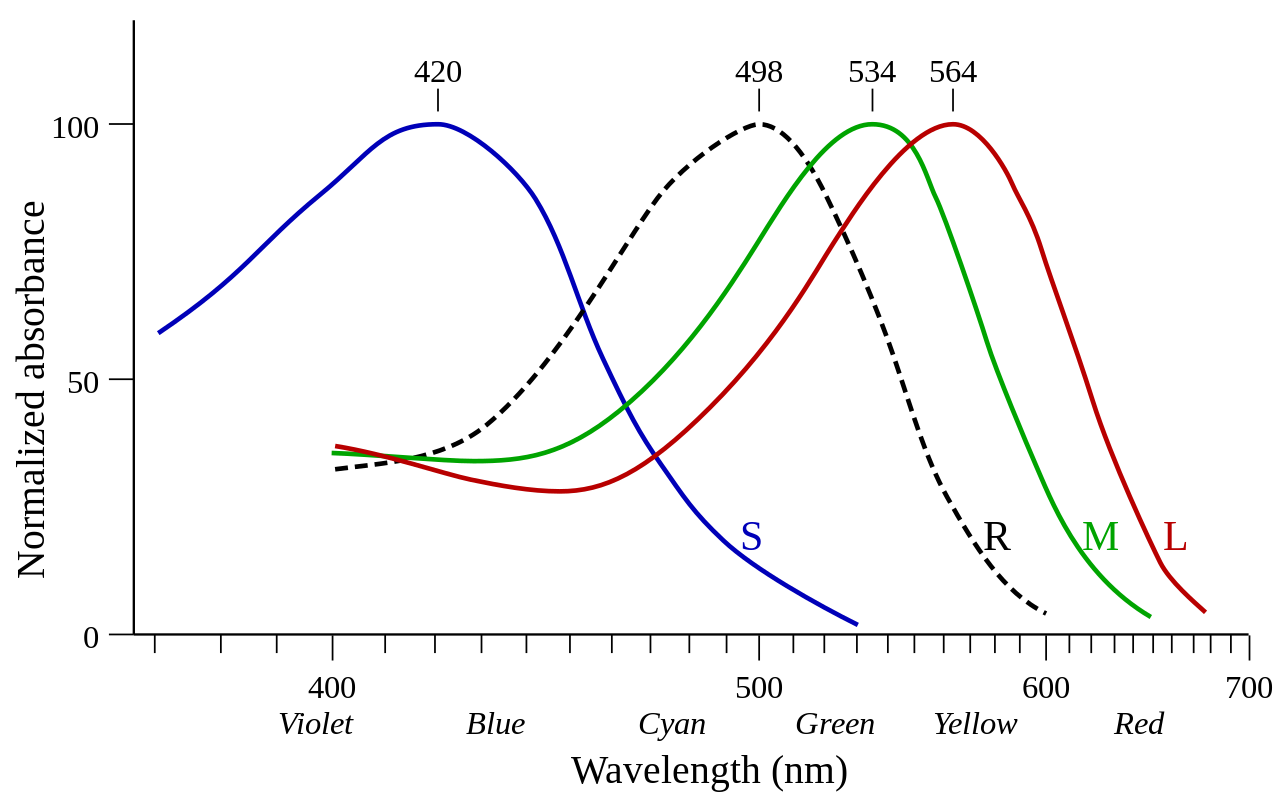
Las opsinas expresadas en las células de los conos se denominan opsinas de cono. [1] Las opsinas de cono se denominan fotopsinas cuando no están unidas al retinal y yodopsinas cuando están unidas al retinal. [1] Las opsinas de cono median la visión fotópica (luz del día). Las opsinas de cono se subdividen según la sensibilidad espectral de su yodopsina, es decir, la longitud de onda en la que se observa la mayor absorción de luz ( λ max ). [7]
| Nombre | Abr. | Celúla | λmáx (nm ) | Variante humana [5] |
|---|---|---|---|---|
| Sensible a las ondas largas | LWS | Cono | 500–570 | OPN1LW eritrolabio "rojo" (564 nm) OPN1MW clorolabino "verde" (534 nm) |
| Sensible a ondas cortas 1 | SWS1 | Cono | 355–445 | OPN1SW cianolabio "azul" (420 nm) (extinto en monotremas ) |
| Sensible a ondas cortas 2 | SWS2 | Cono | 400–470 | (extinto en los mamíferos terios ) |
| Similar a la rodopsina 2 | Rh2 | Cono | 480–530 | (Extinto en mamíferos) |
Opsinas de varilla
Las opsinas expresadas en las células de los bastones se denominan opsinas de bastones. Las opsinas de bastones se denominan escotopsinas cuando no están unidas al retinal y rodopsinas o porfiropsinas cuando están unidas al retinal (1 y 2, respectivamente). Las opsinas de bastones median la visión escotópica (luz tenue). [8] En comparación con las opsinas de los conos, la sensibilidad espectral de la rodopsina es bastante estable y no se desvía mucho de los 500 nm en ningún vertebrado.
| Nombre | Abr. | Celúla | λmáx (nm ) | Variante humana [5] |
|---|---|---|---|---|
| Escotopsina | Rh1 | Vara | Rodopsina: ~500 Porfiropsina: ~522 [3] | Rodopsina humana RHO (498 nm) |
Evolución
| ||||||||||||||||||||||||||||
Los vertebrados actuales suelen tener cuatro clases de opsinas de cono (LWS, SWS1, SWS2 y Rh2), así como una clase de opsinas de bastón (rodopsina, Rh1), todas las cuales fueron heredadas de ancestros vertebrados tempranos. Estas cinco clases de opsinas visuales de vertebrados surgieron a través de una serie de duplicaciones genéticas comenzando con LWS y terminando con Rh1, según el cladograma de la derecha; esto sirve como un ejemplo de neofuncionalización . Desde entonces, cada clase ha evolucionado en numerosas variantes. [9] [10] Las relaciones evolutivas, deducidas utilizando la secuencia de aminoácidos de las opsinas, se utilizan con frecuencia para categorizar las opsinas de cono en su clase respectiva. [1] Los mamíferos perdieron las clases Rh2 y SWS2 durante el cuello de botella nocturno . Los ancestros de los primates desarrollaron más tarde dos opsinas LWS (LWS y MWS), dejando a los humanos con 4 opsinas visuales en 3 clases.
Historia
George Wald recibió el Premio Nobel de Fisiología o Medicina en 1967 por sus experimentos en la década de 1950 que demostraron la diferencia en la absorbancia de estas fotopsinas (ver imagen). [11]
Véase también
Referencias
- ^ abcd Terakita A (1 de marzo de 2005). "Las opsinas". Genome Biology . 6 (3): 213. doi : 10.1186/gb-2005-6-3-213 . PMC 1088937 . PMID 15774036.
- ^ Fasick, Jeffry I.; Robinson, Phyllis R. (23 de junio de 2016). "Adaptaciones de los pigmentos retinianos de los cetáceos a los ambientes acuáticos". Frontiers in Ecology and Evolution . 4 . doi : 10.3389/fevo.2016.00070 .
- ^ de George Wald (1939): El sistema visual de la porfiropsina. En: The Journal of General Physiology . Vol. 22, págs. 775–794. PDF
- ^ Andrew TC Tsin y Janie M. Flores (1985): Regeneración in vivo de la rodopsina y la porfiropsina de los peces de colores. En: J. Exp. Biol. Bd. 122, págs. 269-275. PMID 3723071 PDF
- ^ abc Bowmaker, JK; Dartnall, HJ (1 de enero de 1980). "Pigmentos visuales de bastones y conos en una retina humana". The Journal of Physiology . 298 (1): 501–511. doi :10.1113/jphysiol.1980.sp013097. PMC 1279132 . PMID 7359434.
- ^ Stockman, Andrew; Sharpe, Lindsay T. (junio de 2000). "Las sensibilidades espectrales de los conos sensibles a las longitudes de onda media y larga derivadas de mediciones en observadores de genotipo conocido". Vision Research . 40 (13): 1711–1737. doi :10.1016/S0042-6989(00)00021-3. PMID 10814758. S2CID 7886523.
Como lo deja claro la Fig. 11a, la MSP es de poca utilidad para definir las sensibilidades espectrales de los conos, excepto cerca del fotopigmento λmax. Las grandes discrepancias entre la MSP y otras estimaciones de las sensibilidades espectrales de los conos surgen debido a la pequeña relación señal/ruido de las mediciones de la MSP.
- ^ Gurevich, VV; Gurevich, EV (1 de enero de 2010), Dartt, Darlene A. (ed.), "Fototransducción: inactivación en conos", Enciclopedia del ojo , Oxford: Academic Press, págs. 370–374, doi :10.1016/b978-0-12-374203-2.00190-1, ISBN 978-0-12-374203-2, consultado el 2 de mayo de 2024
- ^ Shichida Y, Matsuyama T (octubre de 2009). "Evolución de las opsinas y la fototransducción". Philosophical Transactions of the Royal Society of London. Serie B, Biological Sciences . 364 (1531): 2881–2895. doi :10.1098/rstb.2009.0051. PMC 2781858 . PMID 19720651.
- ^ Hunt DM, Carvalho LS, Cowing JA, Davies WL (octubre de 2009). "Evolución y ajuste espectral de los pigmentos visuales en aves y mamíferos". Philosophical Transactions of the Royal Society of London. Serie B, Ciencias Biológicas . 364 (1531): 2941–2955. doi :10.1098/rstb.2009.0044. PMC 2781856 . PMID 19720655.
- ^ Trezise AE, Collin SP (octubre de 2005). "Opsinas: evolución en espera". Current Biology . 15 (19): R794–R796. Bibcode :2005CBio...15.R794T. doi : 10.1016/j.cub.2005.09.025 . PMID 16213808.
- ^ Fundación Nobel. «El Premio Nobel de Fisiología o Medicina 1967». Nobelprize.org . Nobel Media AB 2014. Consultado el 12 de diciembre de 2015 .
